醫學原蟲
醫學原蟲
醫學原蟲是寄生在人體管腔、體液、組織或細胞內的致病及非致病性原蟲,約40餘種。其中的一些種類以其獨特的生物學和傳播規律危害人群或家畜,構成廣泛的區域性流行。
原蟲的結構符合單個動物細胞的基本構造,由胞膜、胞質和胞核組成。
包裹蟲體,也稱表膜或質膜。電鏡下可見為一層或一層以上的單位膜結構,其外層的類脂和蛋白分子結合 多糖分子形成表被,或稱糖萼(glycocalyx)。表膜內層可有緊貼的微管和微絲支撐,使蟲體保持一定形狀。研究表明,原蟲的表膜作為與宿主和外環境直接接觸的界面,對保持蟲體的自身穩定和參與宿主的相互作用起著重要的作用。已有證明某些寄生原蟲的表膜帶有多種受體、抗原、酶類,甚至毒素;表膜還具有不斷更新的特點,一些種類的表膜抗原還可不斷變異;在不利條件下,有些種類還可在表膜之外形成堅韌的保護性壁。因此原蟲表膜的功能除具有分隔與溝通作用外,還可以其動態結構參與營養、排泄、運動、感覺、侵襲、隱匿等多種生理活動。對原蟲表膜的深入研究已成為揭示宿主與寄生蟲相互作用機制的重要方面。
主要由基質、細胞器和內含物組成。
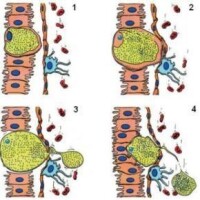
醫學原蟲
原蟲的細胞器按功能分為: ①膜質細胞器:主要由胞膜分化而成,包括線粒體,高爾基複合體,內質網,溶酶體等,大多參與與合成代謝。某些細胞器可因蟲種的代謝特點而有所缺如或獨有,如營厭氧代謝的種類一般缺線粒體;②運動細胞器:為原蟲分類的重要標誌,按性狀分為無定形的偽足(pseudopodium),細長的鞭毛(flagellum),短而密的纖毛(cilia)三種。具相應運動細胞器的原蟲分別稱阿米巴、鞭毛蟲(flagellate)和纖毛蟲(ciliate)。鞭毛蟲和纖毛蟲大多還有特殊的運動器,如波動膜(undulating membrane)。吸盤(sucking disc)以及為鞭毛、纖毛提供動能的神經運動裝置(neuro-motor apparatus)。有些鞭毛蟲的動基體(kinetoplast)即是一種含DNA的特殊細胞器,其功能近似一個巨大的線粒體,含有與之相似的酶。動基體DNA的質和量均與胞核者不同,一些種類已被深入研究用於分子克隆抗體;③營養細胞器:部分原蟲擁有胞口、胞咽、胞肝等幫助取食、排廢。寄生性纖毛蟲大多有伸縮泡能調節蟲體內的滲透壓。此外,鞭毛蟲的胞質可有硬蛋白組成的軸柱(axone),為支撐細胞器,使蟲體構成特定的形態。
原蟲胞質內有時可見多種內含物,包括各種食物泡,營養貯存小體(澱粉泡、擬染色體等),代謝產物(色素等)和共生物(病毒顆粒)等。特殊的內含物也可作為蟲種的鑒別標誌。
為原蟲得以生存、繁衍的主要構造。由核膜、核質、核仁和染色質組成。核膜為兩層單位膜,具微孔溝通核內外。染色質和核仁分別富含DNA和RNA,能被深染。在光鏡下,原蟲胞核需經染色才能辨認,並各具特徵。寄生人體的原蟲多數為泡狀核型(vesicular nucleus),以染色質少而呈粒狀,分佈於核質或核膜內緣和只含一個粒狀核仁為特點。當數纖毛蟲為實質核型(compact nucleus),特點為核大而不規則,染色質豐富,常具一個以上的核仁,故核深染而不易辨認內部。原蟲的營養期大多隻含一個核,少數可有兩個或更多。一般僅在核分裂期核染色質才濃集為染色體,展示染色體核型的形態學特徵。經染色后的細胞核形態特徵是醫學原蟲病原學診斷的重要依據。
原蟲是微波的個體,由於科學技術的發展,醫學原蟲的形態學已深入亞細胞和分子領域。過去在光鏡下未能解決的問題,現可通過超微技術,免疫生化等方法加以判別,從分子水平重新認識。如利什曼原蟲的種群分類,以往難於從光鏡下進行形態學鑒別,今天已可藉助染色體核型、核酸序列構成、酶譜型(zymodeme)或血清學譜型(serodeme)等的綜合分析,達到種群乃至株系的判定。
多數原蟲借運動細胞器進行移位、攝食、防衛等活動。運動方式有偽足運動,鞭毛運動和纖毛運動。沒有細胞器的原蟲也可藉助體表構造進行滑動和小範圍扭轉。具有運動、攝食能力和生殖的原蟲生活史期統稱為滋養體(trophozoite)期,是多數寄生原蟲的基本生活型許多原蟲的滋養體在不良條件下分泌外壁,形成不活動的包囊(cyst)或卵囊(oocyst),用以抵抗不良環境,實現宿主轉換,成為傳播上的重要環節。
寄生原蟲生活在富有營養的宿主內環境,一般可通過表膜以滲透和多種擴散機制吸收小分子養料。多數原蟲還需以細胞器攝食大分子物質,主要有偽足攝食和胞口攝食二種形式。前者有吞噬(phagocytosis)和吞飲(pinocytosis),分別指攝取固態和液態食物,統稱為內胞噬(endocytosis)。纖毛蟲的胞口已早為人知,近代超微研究發現在孢子蟲和鞭毛蟲均有微胞口(micropore)或管胞口(tubular cytostome)等攝食細胞器。攝入的食物在胞質形成食物泡,溶酶體與食物泡結合,參與消化、分解。殘渣和代謝最終產物各以特定的方式,或從胞肛,或從體表,或通過增殖過程的母體裂解而排放於寄生部位。
原蟲的能量代謝和合成代謝,大體符合總論中提到的寄生蟲代謝的一般特徵,但各種群的具體代謝途徑和最終產物則因寄生環境和代謝酶系遺傳性狀的不同而有顯著差異。對於只存在於個別種類中的特殊代謝系統已成為探索合理抗蟲化療方法的研究標靶。已有的研究證明原蟲酶譜的種群間差異與宿主特異性有一定因果關係,酶譜型的分析可能有助於區別某些種類的致病與非致病種群。由於快速增殖,寄生原蟲對蛋白質和多種氨基酸的需求量較多。構成原蟲蛋白的氨基酸種類大多從宿主提供的周圍環境攝入,少數須自身合成。蛋白質的合成在核蛋白體內進行,且極為旺盛,而在通常情況下,蛋白質的分解代謝不佔優勢。有些寄生原蟲的發育增殖往往還需要一些特殊的生長因素或輔助因子,如溶組織內阿米巴及陰道毛滴蟲需要膽固醇;瘧原蟲要求對氨基苯甲酸(PABA)等。
寄生原蟲以無性或有性或兩者兼有的生殖方式增殖,同時以一定的方式排離和轉換宿主以維持種群世代的延續。無性生殖有 ①二分裂:為寄生原蟲最常見的增殖方式,分裂時胞核先分裂,隨後縱向或橫向分裂為二個子體。②多分裂:胞核多次分裂后胞質包繞每個核周圍,一次分裂為多個子代。多分裂形式多樣,瘧原蟲的裂體增殖(schizogony),孢子增殖(sporogony)和某些阿米巴、鞭毛蟲的囊后增殖等都是。③出芽生殖:為大小不等的分裂,如弓形蟲滋養體的內二殖或內二芽殖(endodygony)。有性生殖則可分為接合生殖(conjugation):兩個形態相同的原蟲接合在一起,交換核質後分開各自分裂,多見於纖毛蟲;或配子生殖(Gametogony):先分化為雌雄配子(gamete),而後結合為合子(zygote),再進行無性增殖。配子生殖常為寄生原蟲有性世代的主要階段,本身並無個體增加,卻為無性孢子生殖的先導,如瘧原蟲在蚊體內的發育期。
寄生原蟲的增殖本質上是一種種族生存適應,必然伴隨著排離和宿主更迭。因此醫學原蟲的史完成是從宿
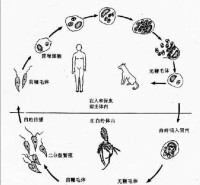
醫學原蟲生活史
1.人際傳播型 生活史只需要一種宿主,憑藉接觸或中間媒介而在人群中直接傳播。可分二類:
⑴生活史只有滋養體階段,以二分裂增殖,直接或間接接觸滋養體而傳播。陰道毛滴蟲,口腔毛滴蟲和齒齦阿米巴等屬此類。
⑵生活史有滋養體和包囊二個階段,前者以二分裂增殖,包囊可有或無核分裂,為有效的排離和傳播階段。多數腸道寄生阿米巴、鞭毛蟲和纖毛蟲屬此類型。
3.蟲媒傳播型 完成生活史需經吸血昆蟲體內的無性或有性繁殖,再接種人體或其它動物。如利什曼原蟲(無世代交替)和瘧原蟲(有世代交替)的生活史。
根據運動細胞器的有無和類型分為鞭毛蟲、阿米巴、纖毛蟲和孢子蟲四大類,生物學分類隸屬於原生生物界(Kingdom Protista),原生動物亞界(Subkingdom Protozoa)之下的三個門,即肉足鞭毛門(Phylum Sarcomastigophora),如動鞭綱、葉足綱;頂復門(Phylum Apicomplexa),如孢子綱;纖毛門(Phylum Ciliophora),如動基裂綱。
| 主要寄生部位 | 蟲名 | 科(Family) | 目(Order) | 綱(Class) |
| 單核吞噬系統 | Leishmania donouani 熱帶利什曼原蟲 Leishmania tropica 巴西利什曼原蟲 Leishmania braziliensis | 錐蟲科 Trypanosomatidae | 動基體目 Kinetoplastida | 動鞭綱 Zoomastigophora |
| 血液 | 錐蟲 Trypanosoma sp. | |||
| 泌尿生殖道 | 陰道毛滴蟲 Trichomonas uaginalis | 毛滴蟲科 Trichomonadidae | 毛滴蟲目 Trichomonadida | |
| 口腔 | 口腔毛滴蟲 Trichomonas tenax | |||
| 腸 | 人毛滴蟲 Trichomonas hominis 脆雙核阿米巴 Dientamoeba fragilis | |||
藍氏賈第鞭毛蟲 Giardia lamblia | 六鞭毛料 Hexamitidae | 雙滴蟲目 Diplomonadida | ||
梅氏唇鞭毛蟲 Chilomastix mesnili | 曲滴蟲科 Retortamonadidae | 旋滴蟲目 Retortamonadida | ||
溶組織內阿米巴 Entamoeba histolytica 哈門氏阿米巴 Entamoeba hartmani 結腸內阿米巴 Entamoeba coli 布氏嗜碘阿米巴 Iodamoeba butschlii 微小內蜒阿米巴 Endolimax nana | 內阿米巴科 Entamoebidae | 阿米巴目 Amoebida | 葉足綱 Lobosea | |
| 口腔 | 齒齦內阿米巴 Entamoeba gingivalis | |||
| 腦(等) | 棘阿米巴 Acanthamoeba sp. | 棘阿米巴科 Acanthamoebidae | ||
福氏耐格里阿米巴 Naegleria fowleri | 雙鞭阿米巴科 Dimastiamoebidiae | |||
| 紅細胞 | 間日瘧原蟲 Plasmodium vivax 三日瘧原蟲 Plasmodium malariae 惡性瘧原蟲 Plasmodium falciparum 卵形瘧原蟲 Plasmodium ovale | 瘧原蟲科 Plasmodidae | 真球蟲目 Eucoccidiida | 孢子綱 Sporozose |
巴貝蟲 Babesia sp. | 巴貝蟲科 Babesidae | 梨漿蟲目 Piroplasmida | ||
| 肺泡 | 卡氏肺孢子蟲 Pneumocystis carinii | 未定 | 真球蟲目 Eucoccidiida | |
| 有核細胞 | 剛地弓形蟲 Toxoplasma gondii | 弓形蟲科 Toxoplasmatidae | ||
| 組織 | 肉孢子蟲 Sarcocystis sp. | 肉孢子蟲科 Sarcocystidae | ||
等孢子蟲 Isospora sp. | 愛美蟲科 Eimeriidae | |||
隱孢子蟲 Cryptosporidium sp. | 隱孢子蟲科 Cryptosporidae | |||
| 結腸 | 結腸小袋纖毛蟲 Balantidium coli | 小袋科 Balantidiidae | 毛口目 Trichostomatida | 動基裂綱 Kinetofragminophorea |
對人體致病的原蟲絕大多數為寄生性,其危害程度因蟲種、株系、寄生部位以及宿主生理狀態而有很大差別。原蟲感染的致病作用,除生物病原因侵襲力與宿主應答水平之間相互作用而導致的機械、化學和生物性質的一般損傷外,還有某些自身的特點。
醫學原蟲
寄生原蟲的微小個體和快速增殖特點,使其致病作用具有與生物病原相似的某種播散潛能。多數致病原蟲 在建立原發病灶后都發現有向近鄰或遠方組織侵蝕和播散的傾向,從而累及多個器官。近代研究已發現致病原蟲具多種利於擴散的因子和生態特點。如原蟲在血細胞內寄生,不僅成為逃避宿主免疫攻擊的一種有效屏障,且為血源播散提供運載工具;利什曼原蟲和弓形蟲被巨噬細胞吞噬后的本特性,使它們能在宿主的免疫活性細胞內增殖自如,並被帶至全身各處,引起累及全身的嚴重感染。近年來在不少致病原蟲與宿主細胞之間發現表面受體作用,這是揭示蟲體對親和細胞或組織進行識別、粘附,進而入侵或噬蝕的物質基礎;溶組織內阿米巴滋養體具有多種膜結合的蛋白水解酶,使它具有接觸溶解宿主組織、細胞的侵襲特性,為其入侵腸壁深層組織,實現實行播散,誘發腸外阿米巴病創造基本條件。應該看到,致病原蟲的播散能力,在致病的傳播上都有重要作用。
臨床發現在一些極度營養不良,晚期腫瘤,長期應用激素製劑及免疫缺陷、免疫功能低下或獲得性免疫缺乏綜合征(艾滋病)患者等常併發致死的原蟲感染。此種因疾病、治療等種種人為或自然因素,招致機體免疫機制削弱而激活某種感染的個體稱為免疫功能受累宿主(immune compromised host)。在寄生原蟲中,有些種群對健康病原蟲(opportunistic pathogen)。常見的有弓形蟲、肺孢子蟲、賈第蟲、隱孢子蟲等。例如有報導晚期艾滋病患者60%合併肺孢子蟲肺炎,成為患者的直接死因;多數表現為隱性感染的弓形蟲病常在白血病及其它惡性腫瘤的治程中急性復燃。條件致病也可導致原蟲對異常部位的侵襲,曾報導一例網織細胞肉瘤患者併發罕見的原發性胃粘膜阿米巴病。
細胞內寄生的原蟲如瘧原蟲、利什曼原蟲,可以逃避宿主抗體的殺傷。有些在血液中的寄生原蟲如錐蟲等,能定期更換體表的抗原(變異),使已產生的特異性免疫失敗。有些寄生原蟲可以誘發對宿主B細胞的多克隆刺激,或抑制宿主的細胞免疫功能,使宿主的特異性免疫能力降低。由於原蟲的增殖作用以及在宿主體內並無成蟲或幼蟲、童蟲等世代之分,被激發的免疫力與蠕蟲引起伴隨免疫有所不同,主要表現為把原蟲抑制在低密度水平的非消除性帶蟲狀態,而出現遷延、反覆和隱性的疾病過程。一些常見的重要原蟲病如瘧疾、阿米巴病、弓形蟲感染等都有這種特殊的臨床表現。能清除蟲體面獲得自愈並對再感染產生完全抵禦力的寄生蟲感染,僅在個別寄生於單核吞噬細胞系統的蟲種如利什曼原蟲中見到。
體積微小,微米計算單細胞結構,功能完善。
