植物水分關係
植物水分關係
植物水分關係,也稱植物水分代謝,植物在生命活動中吸收、傳導和散失水分的連續過程。
植物生命活動中吸收、傳導和散失水分的連續過程,這種關係也稱植物水分代謝。水是植物體的重要組成成分。植物鮮重的70~90%是水。生理活性強的器官如嫩葉、嫩果等的含水量高;而生理活性弱的器官如種子、孢子等則含水較少。細胞中的水分為許多生化反應提供了良好的介質。它的物理性質如高的汽化熱、比熱,對調節植物體溫有重要作用。水也是調節原生質膠體的凝膠與溶膠態的逆變的決定性因素。水還可維持細胞處於緊張狀態,使植株挺立、葉片開展,有利於承受陽光進行光合作用。許多大分子物質的合成也都離不開水。
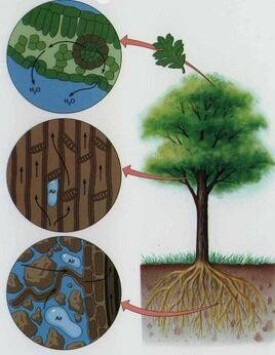
陸生植物由於葉的蒸騰作用,根部從土壤吸收的水,經體內傳導而不斷地向大氣中散失。因此它們必須不斷地在吸水-傳導-散失的連續運動過程中求得體內含水量的動態平衡,才能保證正常的生命活動。水在土壤-植物-大氣系統中的這種運動是由於水在這一系統各個部位中的自由能差所造成的。植物生理學上常用水勢(符號Ψ)表示單位體積(1克分子體積或1毫升水)內水的自由能差,並假定在標準條件下純水的水勢為零值,即單位體積純水的自由能最大。水總是由高自由能區向低自由能區移動。
細胞吸水 細胞水勢主要由溶質勢(ψs)、襯質勢(ψm)和壓力勢(ψp)組成。其中,溶質勢為負值,因溶質溶解使水勢降低;襯質勢也為負值,因構成細胞壁的纖維素和構成原生質的蛋白質對水分子的吸收導致水勢降低,但液泡化細胞的襯質勢很小,通常可忽略不計;壓力勢則為正值,因細胞是一個被胞壁包裹的封閉系統,它在吸水時由於靜水壓力增加而引起的膨壓促使細胞水勢提高。因此細胞水勢隨細胞壓力勢增加而增高,隨溶質勢的降低而降低。細胞水勢越低,吸水力越大。當細胞發生質壁分離時,水勢值與溶質勢值相等,吸水力大。而當細胞吸水達充分緊漲時,水勢值為零;這時細胞內外水分處於動態平衡狀態,細胞停止吸水(見圖)。未發育成熟的未液泡化的細胞開始吸水,主要靠構成細胞的膠體物質如果膠質、纖維素、蛋白質等對水的吸脹作用。如風乾種子的萌發開始吸水,就是靠這種作用。由於未液泡化細胞的溶質勢和壓力勢均為零,種子的吸脹力與襯質勢大小相等,符號相反。若種子置於溶質濃度很高的溶液或土壤中,由於外界水勢低於種子內水勢,種子就吸收不到水分,這是在乾旱土壤播種后種子不能萌發的原因。隨著細胞的成熟和液泡的出現,細胞及其周圍構成一個滲透系統。這時細胞能否吸水主要決定於滲透系統中細胞內外的水勢差。只有當細胞水勢比胞外的水勢更低時,水分才能進入細胞,反之,細胞的水分將向外排出。
當有多個細胞連成一串時,只要兩端之間存在有水勢(或水自由能)差時,水分便可依水勢梯度從高水勢向低水勢移動。在土壤-植物-大氣系統中,水勢梯度一般為土壤水勢>根的水勢>葉的水勢>大氣水勢,水分即沿此梯度而運動。同一器官不同部位細胞之間的水勢也不相同。如根部皮層內部細胞的水勢就低於外部細胞,所以水分才由外向內流動。細胞水勢還受環境因素的影響。在乾旱的情況下,細胞水勢明顯下降,因而也可利用作物葉的水勢作為判斷作物體內水分狀況的指標。
根對水分的吸收根毛區是根的主要吸水部位。其吸收水分的方式分主動吸收和被動吸收兩種。①主動吸收。其特點是水的吸收能逆水勢梯度進行。它是一個生理過程,尤其與呼吸作用關係密切。用生長素類物質如吲哚乙酸、萘乙酸等處理根可改變細胞壁的伸展性,降低壓力勢,從而有促進吸水的作用。根壓也被認為是主動吸水的一種動力,把植物莖稈剪斷後在傷口處有液汁外溢,這種現象稱傷流。傷流是根壓的一種表現,推動傷流的壓力常可有 1~2巴,高的可達5巴。②被動吸收。吸水動力來自蒸騰拉力,它是由葉肉細胞向大氣中散失水分而產生的依次向鄰近細胞取得水分的吸水力。從土壤到葉或從土壤到大氣的水勢梯度是逐漸降低的,因此,只要葉面不斷地蒸騰失水,隨著葉細胞水勢的降低,土壤水分即可沿水勢遞減的順序由根→莖→葉直至散失於大氣之中。
主動吸收與被動吸收在根系吸水中所佔的比重因生育時期與環境條件而異。春天新葉尚未發生或葉子尚未展開前,蒸騰速率低,以主動吸收為主。以後隨著葉片的成長展開而造成蒸騰面積擴大和由於氣溫升高、蒸騰需求增加而造成蒸騰速率提高,被動吸收遂成為根系吸水的主要方式。
水分散失 植物有兩種散失水分的方式:一是以液態散失,稱吐水作用,一是以氣態散失,稱蒸騰作用。植物所吸收的水分95%以上都是通過蒸騰散失的。蒸騰作用是水分子從蒸汽壓較高的葉肉細胞表面向蒸汽壓較低的葉表面大氣擴散的過程。
蒸騰所散失的水分主要是通過氣孔向外擴散的,稱氣孔蒸騰。水分也可通過角質層向外散失,稱角質層蒸騰。二者在蒸騰中所佔比重因生態條件和葉片年齡而異。生長在潮濕地方的植物或陰生植物,或角質層較薄的幼葉片,其角質蒸騰的量大,有時可達總蒸騰量的1/3左右。但一般成熟葉片葉面蒸騰的90%以上是通過氣孔蒸騰。蒸騰速率常被用作蒸騰量的指標,一般以每小時每平方分米葉面蒸騰失水的克數表示。通常白天蒸騰速率約為0.5~2.5克/(分米2·小時),晚上為0.1克以下。
氣孔蒸騰速率受下列關係的調節:因而影響葉內外蒸汽壓力差的環境因素,如光照、溫度、濕度和影響擴散阻力的風速等都可影響蒸騰速率。植物葉片上氣孔的數目、大小及其開張程度影響氣孔阻力的幅度很大,從而顯著影響蒸騰速率。雙子葉植物的氣孔由兩個腎形的保衛細胞組成,靠氣孔一側的內壁厚,而背氣孔的外壁薄。單子葉植物的保衛細胞呈啞鈴形,中間部的胞壁厚,兩頭薄。氣孔的開張是由於保衛細胞吸水膨脹時,厚的細胞壁抗膨脹力強,而薄的細胞壁易於伸長、膨大,因而造成保衛細胞彎曲,使氣孔張開;當保衛細胞失水而體積縮小時,胞壁復原,氣孔關閉。但保衛細胞壓力變化的機理迄今尚未得到充分闡明。主要觀點是認為與光照或黑暗條件下保衛細胞內溶質勢和壓力勢的變化導致細胞水勢的變化有關。保衛細胞內的鉀離子濃度對其水勢變化具有調節作用。
水分傳導 水在植物體內傳導的途徑為:根毛→根皮層→內皮層→根木質部導管→莖木質部導管→葉脈導管→葉肉細胞→氣孔或角質層,然後向大氣散失。只要大氣濕度未達飽和狀態,隨著葉面失水和葉肉細胞水勢降低,按水分由高水勢向低水勢移動的規律,葉肉細胞即可從鄰近水勢較高的細胞,依次向下直至根的表面吸水。一般供水良好的土壤水勢較高,約在0.3~15巴之間,有時接近零。水勢遞減梯度的形成與植株高度相適應,樹冠頂端和上部的葉子要較下部葉子的水勢低。
植物體內水分傳導過程中也存在各種阻力,如進出細胞時穿膜的阻力、各種襯質(細胞壁及其他膠體顆粒等)的阻力、根表面土壤水勢和蒸騰葉的水勢等。阻力越小,則水流量越大。克服水液流動阻力的動力主要來自蒸騰拉力,蒸騰強時細胞失水多,產生的拉力也強。蒸騰拉力的效應與水分子之間相互吸引的內聚力有關,由於後者的存在才使水分子形成一個連續水柱。植物導管內水分子的內聚力可達200巴以上;而體內水柱因受到葉面蒸騰拉力與重力而形成的張力僅為-5~-30巴,水的內聚力遠大於張力。因而水柱受牽引時不易拉斷,水流無須克服很大阻力,即可由下而上。
土壤有效水 多數陸生植物所需水分主要來自土壤。但並非土壤中所有的水分都是植物可利用的。不受重力影響而被土壤保持的水分稱田間持水量,是通常植物可利用水的上限。植物出現永久萎蔫時的土壤含水率稱萎蔫係數或永久萎蔫百分數 (PWP),是植物可利用水的下限。高於萎蔫係數和低于田間持水量的土壤水分屬於對植物有效的水分。水分適當時,由於輸送阻力較小,葉與根無需很低的水勢即可吸進水分。隨著植物對水分的消耗,土壤含水量逐漸降低,阻力增高,根與葉的水勢就需相應降低,才能保持土壤與植物之間有足夠的水勢差,使水分能被植物吸收。當土壤水降低到萎蔫係數時,土壤中已缺乏植物可利用的有效水。因此農業上需及時通過測定土壤水勢變化來確定土壤有效水的含量,為合理排灌提供依據。
水分臨界期 雖然在植物生長發育的任何時期水都是必需的,但不同時期水分的缺乏對植物的影響不同。缺水對植物為害最大的生育階段稱為水分臨界期。各種植物的水分臨界期不盡相同,一般在四分孢子分裂期前後。因此時正值細胞分裂、分化和生長的旺盛時期,蛋白質合成和光合作用等的進行,都對缺水高度敏感。研究植物在一生中的需水規律,確定水分臨界期並按需供水,對確保作物生產和經濟用水都具有十分重要的意義。
植物對水分的需求可根據形態指標,如葉片捲縮、葉色轉濃綠、嫩芽變紅、葉子變韌等判斷。這種方法簡便易行,但不很準確。各種生理指標如細胞水勢和氣孔開度等則可較為準確地說明植物體內的水分平衡狀況。
基本信息
- 中文名
- 植物水分關係
- 別名
- 植物水分代謝
- 拼音
- zhí wù shuǐ fèn guān xì
- 釋義
- 植物在生命活動中水分運動的過程
- 屬性
- 生物生命運動