共找到4條詞條名為細胞分裂的結果 展開
細胞分裂
生物學名詞
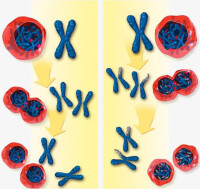
細胞分裂(cell division)是指活細胞增殖其數量由一個細胞分裂為兩個細胞的過程。分裂前的細胞稱母細胞,分裂后形成的新細胞稱子細胞。通常包括細胞核分裂和細胞質分裂兩步。在核分裂過程中母細胞把遺傳物質傳給子細胞。真核細胞分裂包括:有絲分裂、減數分裂、無絲分裂。
核分裂
主要是引發細胞分裂,誘導芽的形成和促進芽的生長。對組織培養的煙草髓或莖切段,細胞分裂素可使已不具備分裂能力的髓細胞重新分裂。這種現象曾被用於細胞分裂素的生物測定。莖切段的分化常受細胞分裂素及生長素比例的調節。當細胞分裂素對生長素的濃度比值高時,可誘導芽的形成;反之則有促進生根的趨勢。如對抑制的腋芽局部施用細胞分裂素或在側芽上塗抹一定濃度的生長素,可以解除頂端對側芽的抑制(即頂端優勢)。天然的簇生植物(蓮座狀植物)或由於病害發生“叢枝病”的植物里,常含有較多的細胞分裂素。
細胞分裂素還有防止離體葉片衰老、保綠的作用,這主要是由於細胞分裂素能夠延緩葉綠素和蛋白質的降解速度,穩定多聚核糖體(蛋白質高速合成的場所),抑制DNA酶、RNA酶及蛋白酶的活性,保持膜的完整性等。在葉片上局部施用細胞分裂素,能吸聚其他部分的物質向施用處運轉和積累。
細胞分裂素的作用方式還不完全清楚。已知在tRNA中與反密碼子相鄰的地方有細胞分裂素,在蛋白質合成過程中,它們參與到tRNA與核糖體mRNA複合體的連接物上。但這可能不是外源細胞分裂素的作用方式。因為在tRNA中,細胞分裂素的合成是由原來在tRNA中的嘌呤的改變產生的。而外源細胞分裂素並不參入tRNA中,但可促進硝酸還原酶、蛋白質和核酸的合成。除了天然的促進細胞分裂的物質外,還用化學方法人工合成了一些類似激動素的物質。通常也統稱細胞分裂素。其中活性較強,也最常用的是6-苄基嘌呤。
還了解不多,只對少數細菌的分裂有些具體認識。原核細胞既無核膜,也無核仁,只有由環狀DNA分子構成核區,又稱擬核,具有類似細胞核的功能。擬核的DNA分子或者連在質膜上,或者連在質膜內陷形成的質膜體上,質膜體又稱間體。隨著DNA的複製間體也複製成兩個。以後,兩個間體由於其間的質膜的生長而逐漸離開,與它們相連接的兩個DNA分子環於是被拉開,每一個DNA環與一個間體相連。在被拉開的兩個DNA環之間細胞膜向中央長入,形成隔膜,終於使一個細胞分為兩個細胞。
按細胞核分裂的狀況可分為3種:即有絲分裂、減數分裂和無絲分裂。有絲分裂是真核細胞分裂的基本形式。減數分裂是在進行有性生殖的生物中導致生殖母細胞中染色體數目減半的分裂過程。它是有絲分裂的一種變形,由相繼的兩次分裂組成。無絲分裂又稱直接分裂。其典型過程是核仁首先伸長,在中間縊裂分開,隨後核也伸長並在中部從一面或兩面向內凹進橫縊,使核變成腎形或啞鈴型,然後斷開一分為二。差不多同時細胞也在中部縊裂分成兩個子細胞,由於在分裂過程中不形成由紡錘絲構成的紡錘體或中心體發出的星射線,不發生由染色質濃縮成染色體的變化,故命名無絲分裂。
無絲分裂時由於不經過染色體有規律的平均分配,故存在遺傳物質不能保證(但也不是沒有可能)均等分配的問題,由此有些人認為這是一種不正常的分裂方式.
無絲分裂是最早發現的一種細胞分裂方式,早在1841年雷馬克(R.Remak)於雞胚血球細胞中見到。在無絲分裂中,核仁、核膜都不消失,沒有染色體的出現,在細胞質中也不形成紡錘體,當然也就看不到染色體複製和平均分配到子細胞中的過程。但進行無絲分裂的細胞,染色體也要進行複製,並且細胞要增大。當細胞核體積增大一倍時,細胞就發生分裂。至於核中的遺傳物質DNA是如何分配到子細胞中的,還有待進一步研究。無絲分裂是最簡單的分裂方式。過去認為無絲分裂主要見於低等生物和高等生物體內的衰老或病態細胞中,但後來發現在動物和植物的正常組織中也比較普遍地存在。無絲分裂在高等生物中主要是高度分化的細胞,在動物的上皮組織、疏鬆結締組織、肌肉組織和肝組織中,在植物各器官的薄壁組織、表皮、生長點和胚乳等細胞中,都曾見到過無絲分裂現象。
分裂生殖又叫裂殖,是無性生殖中常見的一種方式,即是母體分裂成2個(二分裂)或多個(復分裂)大小形狀相同的新個體的生殖方式。這種生殖方式在單細胞生物中比較普遍,但對不同的單細胞生物來說,在生殖過程中核的分裂方式是有所不同的,可歸納為以下幾種方式:
1以無絲分裂方式營無性分裂生殖
原核細胞的分裂包括兩個方面:(1)細胞DNA的分配,使分裂后的子細胞能得到親代細胞的一整套遺傳物質;(2)胞質分裂把細胞基本上分成兩等分。
複製好的兩個DNA分子與質膜相連,隨著細胞的生長,把兩個DNA分子拉開,細胞分裂時,細胞壁與質膜發生內褶,最終把母細胞分成了大致相等的兩個子細胞。
2以核的有絲分裂方式營無性分裂生殖
(1)甲藻細胞染色體的結構和獨特的有絲分裂,兼有真核細胞和原核細胞的特點,細胞開始分裂時核膜不消失,核內染色體搭在核膜上,分裂時核膜在中部向內收縮形成凹陷的槽,槽內細胞質出現由微管按同一方向排列的類似於紡錘絲的構造,調節核膜和染色體,分離為子細胞核,最終分裂成兩個子細胞(甲藻)。
(2)眼蟲營分裂生殖時,核進行有絲分裂,分裂過程中核膜並不消失,隨著細胞核中部收縮分離成兩個子核,然後細胞由前向後縱裂為二(縱二分裂),其中一個帶有原來的一根鞭毛,另一個又長出一根新鞭毛,從而形成兩個眼蟲。
(3)變形蟲長到一定大小時,進行分裂繁殖,是典型的有絲分裂,核膜消失,隨著細胞核中部收縮,染色體分配到子核中,接著胞質一分為二,將細胞分裂成兩個子代個體。
3以核的無絲分裂和有絲分裂方式營無性分裂生殖
這種方式最典型的代表就是草履蟲,草履蟲屬原生動物纖毛蟲綱,細胞內有大小兩種類型的核,即大核和小核,小核是生殖核,大核是營養核,在草履蟲進行無性繁殖時,小核進行核內有絲分裂,大核則行無絲分裂,接著蟲體從中部橫縊分成2個新個體。
植物細胞通過分裂進行繁殖。繁殖是生物或細胞形成新個體或新細胞的過程。植物細胞的分裂包括無絲分裂、有絲分裂和減數分裂和細胞的自由形成等不同的方式。
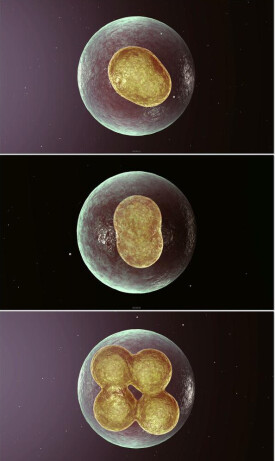
有絲分裂又稱為間接分裂,它是一種最普遍,是常見的分裂方式。有絲分裂為連續分裂,一般分為核分裂和胞質分裂。
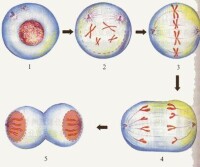
有絲分裂的過程
間期:分為G1,S,G2,主要進行DNA複製和相關蛋白質合成,核膜核仁逐漸消失。
前期:核內的染色質凝縮成染色體,核仁解體徹底消失,核膜破裂以及紡錘體開始形成。
中期:中期是染色體排列到赤道板上,紡錘體完全形成時期。
後期:後期是各個染色體的兩條染色單體分開,在紡錘絲的牽引下,分別由赤道移向細胞兩極的時期。
末期:為形成二子核和胞質分裂的時期。染色體分解,核仁、核膜出現,赤道板位置形成細胞板,將來形成新的細胞壁。赤道板上堆積的紡錘絲,稱為成膜體。
動物細胞與植物細胞相似,不同的是動物細胞是有中心體發出星射線形成紡錘體,植物細胞是從兩級直接發出紡錘絲。有絲分裂末期動物細胞細胞膜向內凹陷,形成兩個子細胞,植物細胞是在赤道板(虛擬想象)位置形成細胞板,將細胞分成兩個子細胞。
2、細胞質分裂(時間短):核分裂後期,染色體接近兩極時,細胞質分裂開始。在兩個子核之間的連續絲中增加了許多短的紡錘絲,形成一個密集著紡錘絲的桶狀區域,稱之為成膜體。微管的數量增加,成膜體中有來自高爾基體和內質網的泡囊(含多糖類物質),沿著微管指引方向,聚集,融合,釋放出多核物質,構成細胞板,從中間開始向周圍擴展,直至與母細胞壁相連,成為胞間層——初生壁,新質膜由泡囊的被膜融合而成。新細胞壁形成后,把兩個新形成的細胞核和它們周圍的細胞質分隔成為兩個子細胞。
3、有絲分裂的特點:通過細胞分裂使每一個母細胞分裂成兩個基本相同的子細胞,子細胞染色體數目、形狀、大小一樣,每一染色單體所含的遺傳信息與母細胞基本相同,使子細胞從母細胞獲得大致相同的遺傳信息。使物種保持比較穩定的染色體組型和遺傳的穩定性。
有性生殖要通過兩性生殖細胞的結合,形成合子,再由合子發育成新個體。生殖細胞中的染色體數目是體細胞中的一半。(否則生物每繁殖一代,體細胞中的染色體數目就會增加一倍)。既然在形成生殖細胞——精子或卵細胞時,染色體數目要減少一半,則原細胞必須經過減數分裂。
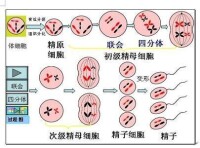
(一)精子形成
精子的形成部位:睾丸(精巢)的曲細精管中。在精巢中,通過有絲分裂產生了大量的原始生殖細胞,也就是精原細胞。根據有絲分裂的特徵,可知精原細胞的染色體數目與體細胞染色體數目是相同的。在精原細胞時期,進行了染色體複製。當雄性動物性成熟后,睾丸里的一部分精原細胞就開始進行減數分裂,經過減數分裂后,形成精細胞,精細胞經過變性形成男性生殖細胞——精子
精原細胞在減數分裂過程中連續進行了兩次分裂
1間前
1間后(複製)
精子形成過程
1中期1後期
減數分裂
2前期2中期
2後期2末期
3變形期
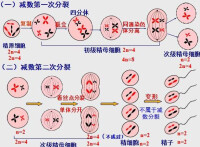
第一次分裂四分體分離,染色體數目減半;
第二次分裂兩條姐妹染色單體分離,染色體數目不變。
第一次分裂的前期,細胞中的同源染色體兩兩配對,叫聯會。所謂的同源染色體,指減數分裂時配對的兩條染色體,形狀和大小一般都相同,一個來自父方,一個來自母方。聯會後,染色體進一步螺旋化變粗,逐漸在光學顯微鏡下可見每個染色體都含有兩個姐妹染色單體,由一個著絲點相連,每對同源染色體則含有四個姐妹染色單體,叫四分體。把四分體時期和聯會時比較,由於染色體複製在精原細胞時就發生了,因此,它們所含的染色單體、DNA數目都是相同的,不同的主要是染色體的螺旋化程度不同,聯會時染色體螺旋化程度低,染色體細,在光學顯微鏡下還看不清染色單體,因此,沒有在圖上表示出來。四分體時期,染色體螺旋化程度高,染色體變粗了,可在光學顯微鏡下清楚地看到每一個染色體有兩個單體。
在細胞分裂的同時,細胞內的同源染色體彼此分離,結果一個初級精母細胞便分裂成兩個次級精母細胞,而此時細胞內的染色體數目也減少了一半,細胞內不再存在同源染色體。減數第一次分裂結束。
減數第二次分裂是從次級精母細胞開始的,細胞未經染色體的複製,直接進入第二次分裂。在細胞第二次分裂過程中,染色體的行為和前面所學的有絲分裂過程中染色體的行為非常相似,細胞內染色體的著絲點排列在赤道板這一位置后,接著進行分裂,於是兩條姐妹染色單體分離,分別移向細胞兩極。與此同時,細胞分裂,結果生成了精子細胞。精子細胞經過變形后成為精子,兩個次級精母細胞最後生成了四個精子,減數分裂結束。
隨後,各個四分體排列在細胞中央,同源染色體好像手拉手似地排成兩排,紡錘絲收縮,牽引染色體向兩極移動,導致四分體平分為二,配對的同源染色體分開,但此時著絲點並未分開,每一染色體上仍有兩條染色單體。接著發生細胞分裂,一個初級精母細胞分裂成兩個次級精母細胞,而每個次級精母細胞中的染色體數目就只有初級精母細胞的一半了,初級精母細胞有4條染色體,而次級精母細胞只有兩條染色體,染色體數目減半的原因是同源染色體分開,在次級精母細胞中已沒有同源染色體了。
聯會的同源染色體分開,說明染色體具有一定的獨立性,由於兩個同源染色體在細胞中央的排列位置是隨機的,可以互相交換,因此,就決定了同源的兩個染色體各移向哪一極也是隨機的,這樣,不同對的染色體之間就可以自由組合。這是將來要學的基因的自由組合規律的細胞學基礎。
第二次分裂的基本過程與有絲分裂相似:中期,染色體的著絲點排成一排,後期,著絲點一分為二,兩個姐妹染色單體成為兩個染色體,在紡錘絲的牽引下,移向兩極,接著,細胞分裂,兩個次級精母細胞分裂成4個精子細胞,減數分裂完成。
精子細胞再經過變形,形成精子,在這個過程中,丟掉了精子細胞的大部分細胞質,帶上重要的物質——細胞核內的染色體,輕裝上陣,並形成了一個長長的尾,便於遊動。
(二)卵細胞形成
卵細胞在卵巢中形成,其過程與精子形成過程基本相同,但也有區別。相同點:染色體複製一次,都有聯會和四分體時期,經過第一次分裂,同源染色體分開,染色體數目減少一半,在第二次分裂過程中,有著絲點的分裂,最後形成的卵細胞,它的染色體數目也比卵原細胞減少了一半。不同點:每次分裂都形成一大兩小三個細胞,小的叫極體,極體以後都要退化,只剩下一個卵細胞,而一個精原細胞是形成4個精子;卵細胞形成后,不需要經過變形,而精子要經過變形才能形成。卵細胞:細胞體形較大,呈球形,不能遊動;含卵黃多,營養物質豐富,保證受精后發育成新個體。精子:細胞體形較小,有鞭毛,能遊動,其特點是保證受精作用的實現;卵細胞的第二次減數分裂中期是在受精作用完成時才開始發生。
受精作用——精子與卵細胞結合成為合子的過程
精子的頭部進入卵細胞,精子與卵細胞的細胞核結合在一起,因此,合子中染色體數目又恢復到原來的體細胞的數目,其中一半來自精子(父方),一半來自卵細胞(母方)。從同源染色體的角度看,精子和卵細胞中的同源染色體都是成單存在,但精子帶有其中的一條,卵細胞帶有其中的另一條,受精后,這兩條同源染色體到了一個細胞中,它們就成對存在了,所以,關於同源染色體的概念說,一條來自父方,一條來自母方,就是這個意思。
減數分裂使染色體數目減半,受精作用使染色體數目又恢復到原來的數目,從而使生物遺傳後代染色體數目保持恆定。
細菌可以以無性或者遺傳重組二種方式繁殖,最主要的方式是以二分裂這種無性繁殖的方式:一個細菌細胞壁橫向分裂,形成兩個子代細胞。
除細菌以外,二分裂也是原生動物最普遍的一種無性生殖.一般是有絲分裂,分裂時細胞核先由一個分為二個,染色體均等的分佈在兩個子核中,隨後細胞質也分別包圍兩個細胞核,形成兩個大小、形狀相等的子體,二分裂可以是縱裂,如眼蟲;也可以是橫裂,如草履蟲;或者是斜分裂,如角藻。
細菌沒有核膜,只有一個大型的環狀DNA分子,細菌細胞分裂時,DNA分子附著在細胞膜上並複製為二,然後隨著細胞膜的延長,複製而成的兩個DNA分子彼此分開;同時,細胞中部的細胞膜和細胞壁向內生長,形成隔膜,將細胞質分成兩半,形成兩個子細胞,這個過程就被稱為細菌的二分裂。無絲分裂則是發現最早的一種真核細胞的分裂方式,在真核生物中普遍存在,而且不僅在體細胞中,甚至在生殖細胞中都能進行無絲分裂。由於其核分裂的過程不出現染色體和紡錘絲,胞質分裂后的遺傳物質不一定能夠平均分配給子細胞,與有絲分裂有很大區別,故稱無絲分裂。無絲分裂的過程大致可劃分為4個時期:第一期核內染色質複製倍增,核及核仁體積增大,核仁及核仁組織中心分裂。第二期以核仁及核仁組織中心為分裂制動中心,以核仁與核膜周染色質相聯繫的染色質絲為牽引帶,分別牽引著新複製的染色質和原有的染色質。新複製的染色質在對側核仁組織中心發出的染色質絲的牽引下,離開核膜移動到核的赤道面上。第三期核拉長成啞鈴型,中央部分縊縮變細。第四期核膜內陷加深,最終縊裂成為兩個完整的子細胞核;接著,整個細胞從中部縊裂成兩部分,形成兩個子細胞。由此我們不難看出:無絲分裂和二分裂有著本質的區別,二分裂指的是原核生物進行的一種最原始的細胞增殖方式,而無絲分裂是真核生物獨特的細胞增殖方式,通過這種分裂,可同時形成多個核;且分裂時細胞核仍可執行其生理功能。
以高等動物為例,受精卵卵裂進行到一定時間細胞增多,形成了一個內部有腔的球狀胚,這個時期的胚叫囊胚。這時期的胚其特點是中央有一空腔,叫囊胚腔。胚繼續發育形成原腸胚。由於動物極一端的細胞分裂較快,新產生的細胞便向植物極方向推移、使植物極一端的細胞向囊胚腔陷入,囊胚腔縮小,內陷的細胞不僅構成了胚胎的內胚層,而且圍成了一個新的腔叫原腸腔。在內外細胞層之間分化出了一個新的細胞層,叫做中胚層,這時期的胚就叫原腸胚。原腸胚的特點是:具有原腸腔和外、中、內三個胚層。原腸胚的外胚層由包被胚胎表面的動物極一端的細胞構成,內胚層由陷入囊胚腔的細胞構成,中胚層位於內、外胚層之間,這三個胚層繼續發育,經過組織分化、器官形成,最後形成一個完整的幼體。
外胚層:形成神經系統的各個器官,包括腦、脊髓和神經、眼的 網膜、虹膜上皮、內耳上皮、以及皮膚的表皮和皮膚的附屬結構。
內胚層:形成消化道(咽、食道、胃、腸等)和呼吸道(喉、氣管、支氣管等)的上皮,肺、肝、胰和咽部分衍生的腺體(甲狀腺,副甲狀腺、胸腺等)以及泌尿系統的膀胱、尿道和附屬腺體的上皮等。
中胚層:主要形成各種肌肉、骨胳、結締組織以及皮膚的真皮,循環系統(心臟、血管和血液)、排泄系統(腎、輸尿管)、生殖系統(生殖腺、生殖管道及附腺等)、氣管和消化道的管壁、體腔膜等。
細胞分化在胚胎期達到最大限度。
幹細胞(ES)
是一類具有自我更新和分化潛能的細胞。它包括胚胎幹細胞和成體幹細胞。幹細胞的發育受多種內在機制和微環境因素的影響。人類胚胎幹細胞已可成功地在體外培養。最新研究發現,成體幹細胞可以橫向分化為其他類型的細胞和組織,為幹細胞的廣泛應用提供了基礎。
在胚胎的發生髮育中,單個受精卵可以分裂發育為多細胞的組織或器官。在成年動物中,正常的生理代謝或病理損傷也會引起組織或器官的修復再生。胚胎的分化形成和成年組織的再生是幹細胞進一步分化的結果。胚胎幹細胞是全能的,具有分化為幾乎全部組織和器官的能力。
到了個體發育的一定階段甚至成體,仍有一部分細胞負責組織的更新和修復,諸如血液、腸道粘膜上皮、皮膚表皮等。這些細胞便是一般所指的特定組織的幹細胞,又稱為多能性細胞。
隨著細胞生物學的發展,人們現已發現,栽些成體組織不但能再生,而且可以衍生成與其來源不同的細胞類型。例如肌肉細胞在一定的環境下可以成為有增殖能力的骨髓細胞;相反地,血液“前體細胞”(即未完全成熟的血細胞)也可變成肌肉細胞,甚至長出肝或腦細胞來。
ES雖好,但其來源有限。ES多取自人工流產的極早期胚胎或是培植試管嬰兒時剩餘的胚胎。然而現已有科學家證實ES可以在體外即實驗室的試管中培養與繁殖,並且可以使ES細胞增殖、定向分化並形成多巴胺能性細胞,而這正是治療帕金森病所亟需的神經元。
起開關作用的蛋白質名為“GATA”。研究人員利用基因工程方法使老鼠胚胎幹細胞的“GATA”含量增加,結果胚胎幹細胞變成了在孕育生命階段起重要作用的其他細胞。研究人員還同時發現,除了蛋白質“GATA”,還有其他物質也起到開關作用,它們相互合作,共同決定胚胎幹細胞的命運。
研究人員計劃通過基因技術找到所有“開關”,這樣胚胎幹細胞就會按人的意志生成各種組織。操作好這些“開關”,可能使普通幹細胞變成真正的“萬能細胞”。
幹細胞尤其是胚胎幹細胞的識別、分離、增殖、定向分化將成為細胞生物學以及整個生命科學的主攻熱點。
一個新的有趣的發現是,生命細胞活著時左旋,死亡后即右旋,一切病毒細菌和死亡的物質卻只會右旋不會左旋。這是什麼原因?這是否與宇宙本來就是左右不對稱有關(地球的自轉、公轉仍然是左旋的,十大行星幾乎都是左旋的,宇宙大黑洞也是左旋的,中微子現也認為是左旋的)。看來衰老問題聯繫到更廣闊的研究領域。
植物組織培養愈傷組織
花藥培養anther culture用植物組織培養技術,把發育到一定階段的花藥,通過無菌操作技術,接種在人工培養基上,通過植物激素改變花藥內花粉粒的發育程序,誘導其分化,並連續進行有絲分裂,形成細胞團,進而形成一團無定型的無分化的薄壁組織——愈傷組織,再分化成胚狀體,隨後改變激素的比例使愈傷組織分化形成完整的植株。
亦稱癒合組織或創傷組織。植物體局部受傷后,在傷口表面形成的具有分生能力和保護作用的活的薄壁細胞群。愈傷組織的外層細胞常可木質化或形成周皮,對其表層的細胞起保護作用。在植物嫁接中愈傷組織促使砧木與接穗緊密結合,植物扦插能從愈傷組織分化出不定根和不定芽;在組織和細胞培養時,條件適宜也能長出愈傷組織。利用愈傷組織誘導形成新植株已廣泛應用於植物的無性繁殖(即人造種子)。
植物組織培養中愈傷組織的形成和形態發生(植物體的結構層次1課時)
在植物組織培養中,主要目標是誘導愈傷組織形成和形態發生變化,使一個離體的細胞、一塊組織或一個器官的細胞,通過脫分化形成愈傷組織,並由愈傷組織再分化形成植物體。
愈傷組織的形成從一塊外植體形成典型的愈傷組織,大致要經歷三個時期:起動期、分裂期和形成期。
起動期是指細胞準備進行分裂的時期。用於接種的外植體的細胞,通常都是成熟細胞,處在靜止狀態。起動期是通過一些刺激因素(如機械損傷、改變光照強度、增加氧等)和激素的誘導作用,使外植體細胞的合成代謝活動加強,迅速進行蛋白質和核酸的合成。機械損傷能誘導植物體細胞開始分裂,如傷口上會出現愈傷組織。在植物組織培養中沿用了愈傷組織這一名詞,但是植物組織培養中誘導外植體細胞分裂形成的愈傷組織,大都不是損傷的結果。外源的生長素類物質對誘導細胞開始分裂效果很好,因此生長素類物質在植物組織培養中得到了廣泛應用,常用的有2,4—二氯苯氧乙酸、萘乙酸、吲哚乙酸和細胞分裂素等。
分裂期是指外植體細胞經過誘導以後脫分化,不斷分裂、增生子細胞的過程。處於分裂期的愈傷組織的特點是:細胞分裂快,結構疏鬆,顏色淺而透明。
外植體的脫分化因植物種類、器官來源及其生理狀況的不同而有很大差別。例如,煙草、胡蘿蔔等植物的脫分化比較容易,禾本科植物的脫分化比較難;花的脫分化比較容易,莖、葉的脫分化比較難;幼嫩組織的脫分化比較容易,成熟的老組織脫分化比較難。
分化期是指在分裂期的末期,細胞內開始出現一系列形態和生理上的變化,從而使愈傷組織內產生不同形態和功能的細胞。這些細胞類型有薄壁細胞、分生細胞、色素細胞、纖維細胞,等等。
外植體的細胞經過起動、分裂和分化等一系列變化,形成了無序結構的愈傷組織。如果在原來的培養基上繼續培養愈傷組織,會由於培養基中營養不足或有毒代謝物的積累,導致愈傷組織停止生長,甚至老化變黑、死亡。如果要讓愈傷組織繼續生長增殖,必須定期地(如2~4周)將它們分成小塊,接種到新鮮的培養基上,這樣愈傷組織就可以長期保持旺盛的生長。
愈傷組織的形態發生方式 經過起動、分裂和分化期產生的愈傷組織,其中雖然發生了細胞分化,但是並沒有器官發生。只有滿足某些條件,愈傷組織的細胞才會發生再分化,產生芽和根,進而發育成完整植株。愈傷組織的形態發生方式主要有不定芽方式和胚狀體方式兩種。不定芽方式是在某些條件下,愈傷組織中的分生細胞發生分化,形成不同的器官原基,再逐漸形成芽和根。胚狀體方式是由愈傷組織細胞誘導分化出具有胚芽、胚根、胚軸的胚狀結構,進而長成完整植株。這種由愈傷組織中的薄壁細胞不經過有性生殖過程,直接產生類似於胚的結構,叫做胚狀體。
不定芽方式和胚狀體方式是植物組織培養中最常見和最重要的兩種方式。胚狀體方式比不定芽方式有更多的優點,如胚狀體產生的數量比不定芽多,胚狀體可以製成人工種子,等等。
肝細胞分裂
肝臟是人體一個重要的消化器官,因其在損傷的情況下有強大的再生修復能力而一直受到醫學家們的普遍關注。認為參與肝臟修復的細胞可能有三個來源:一是通過肝細胞自身的有絲分裂來彌補死亡的肝細胞,這在正常的肝細胞代謝及輕度的肝臟損傷中起主要作用;二是在比較較嚴重的肝損情況下,肝臟的幹細胞被激活並向肝細胞分化以修復肝臟;最近的研究顯示骨髓中的造血幹細胞也具有向肝細胞分化能力,提示可作為肝臟細胞修復第三個潛在來源1。
影響因素
自然情況下:
內因:細胞周期受到一系列基因、酶和蛋白質等內在因素的精確調控。不同的組織基因選擇性表達會造成差異。
外因:主要受一些外界信號的刺激,有細胞因子(如肽類生長因子)、激素和細胞外基質等。
1.肽類生長因子:主要通過旁分泌方式作用於靶細胞;同時也存在著自分泌,當兩種方式均分泌不足時會抑制增殖與分化。
2.激素:只作用於特定靶細胞促進其生長與分化。
3.細胞外基質:應其特定細胞的增殖與分化。主要通過與細胞表面整聯蛋白作用激活相關酶,啟動相關信號。它對幹細胞的增殖分化具有誘導作用。在發育和創傷組織中,透明質酸合成旺盛,促進細胞的增殖與遷移,抑制細胞分化。當增殖到一定程度時透明質酸會水解。
又如胚胎時期,下調刺激信號的水平,抑制細胞周期引擎會抑制細胞分裂。在肌肉發生過程中,屬於TGF-β超家族的肌肉抑制素是肌肉生長的負調控因子。
非自然情況下,射線、低溫﹑化學藥劑、病毒等環境因素對細胞分裂的影響是通過內因起作用的,即通過導致基因突變或影響酶的活性而影響細胞分裂。
分裂新形式
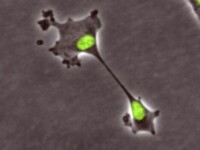
“核分裂”(klerokinesis)
研究小組給人類細胞複製出了多倍染色體,以模擬癌症。他們用一種常規化學物質阻止了胞質分裂,結果發現分裂並未顯出異常。
他們進一步觀察了人類細胞是怎樣恢復正常染色體倍數的。從一個細胞變成兩個核開始觀察,吃驚地發現細胞沒經過有絲分裂,而是直接由一個細胞變成了兩個細胞。每個新細胞都遺傳了一個完整無缺的細胞核,包含一套完整染色體。分裂發生的時間出乎預料,是在延遲生長階段,而不是在有絲分裂結束時。他們還做了大量額外實驗,以確定這種分裂和正常的細胞分裂形式“胞質分裂”不同。
他們還發現,有90%的子細胞恢復為正常的配對染色體。在一個生物經過的所有細胞分裂周期中,每次的胞質分裂偶爾也會失敗。這種新分裂是一種補救機制,讓細胞能從故障中恢復正常。
能夠影響細胞分裂的因素很多,而且極為複雜,目前還沒達到對其全面認識的水平。
1.細胞的表面積與體積之比以及細胞核與細胞質體積之間的平衡:細胞通過它的表面不斷地與周圍環境或鄰近細胞進行物質交換,這麼它就必須有足夠的表面積,否則它的代謝作用就很難進行。但細胞的體積由於生長而逐漸增大時,表面積與體積的比例就會變得越來越小,物質交換適應不了細胞的需要,這可以引起細胞的分裂,以恢復適宜的比例。同樣的,細胞核中的遺傳信息指引和控制範圍有限,細胞核對太大範圍的細胞質的調控作用就會相對減少。曾做過這樣的實驗:當人工培養的變形蟲快要分裂的時候,把它的細胞質切去一大塊,這個變形蟲就不再分裂。等它長大起來又要分裂的時候又切去一塊,它也不再分裂。但如果讓其繼續生長,體積達到一定大小時,它又會分裂起來。
2.抑素
3.cAMP
4.激素
5.接觸抑制(contact inhibition)
以及其它種種因素
在地球生命演化的早期,為何會出現出細胞分裂的特性?有學者提出了細胞分裂的光合起源假說。首先,細胞本質上必須是一個獨立的半開放體系,細胞膜允許物質的進出—營養物質的吸收以及代謝物的輸出,這是生命個體進化的基礎。這樣,細胞膜就必須具有選擇性的通透性,允許一些小分子化合物(如養分)的進出,但是,大的分子肯定是難以自由通行的。
脂雙層膜正好具有了這樣的選擇通透性:水、甘油、色氨酸、一個普通的蛋白質以及鈉離子在卵磷脂膜中的相對滲透性分別為10、10、10、1和1,這就是說,雙層磷脂膜對大分子和帶電離子是不通透的,而小的非帶電分子有較高的通透性,因此,這樣的半透膜建立了對不同類型(如極性、大小)分子選擇性攝取或釋放的體系,而生物大分子若在囊泡內形成,則能在囊泡內部很好地保存并行使生命功能。帶電粒子不能自由穿越脂雙層這一點十分重要,因為如果沒有這一特性,H就不能形成跨膜電位,也就不能用於ATP的合成,更不會有現在生命系統的存在!
接下來,如果在前細胞體中發生了有機物質的合成,就會出現新的問題。在一個有限的前細胞體中如果發生光合作用驅動的有機物合成,就必然會導致有機物質在細胞體中的不斷增加,也必然會導致細胞的破裂,或許正是這種反反覆復的細胞破裂過程(脂質的化學特性決定了它們在水溶液中能自發形成脂雙層的球形結構,這是這種過程能不斷重複的重要基礎)後來演化出生命最重要的機制之一——細胞分裂。因此,能進行光合作用的原始細菌(最初可能沒有產氧能力)應該是最先出現的單細胞生物。
或者簡單地說,為了進行有機物質的合成,細胞膜必須對物質的進出有選擇性,特別是它必須保證養分的進入和一部分廢物的排出。但是,這也不能解決所有問題,譬如,它如何來處置光合作用生產的有機物的堆積問題呢?可以設想有三種可能的途徑:1)細胞不斷增大。但事實上,這是不可能的,因為對漂浮於水中的原始細胞來說,細胞的增大就會由於沉降的加速而使它們快速葬身海底的風險聚增;2)細胞將合成的有機物迅速分解,排出體外;3)隨著有機物的堆積,細胞一分為二,不斷重複新的合成—分裂之過程。顯然,生命選擇了第3種方案。
這種策略似乎與現生細胞體積的周期性變化相吻合:細胞分裂形成的新細胞,最初體積較小,只有母細胞的一半,但它們能迅速合成新原生質,細胞隨之增大,到母細胞一般大小時,便可繼續分裂,如此循環往複。利用光能不斷合成有機物的一個結果就是,要求細胞體積不斷增大,如果偶然出現分裂的細胞,而且有一部分存活下來,而且逐漸被記憶(遺傳)下來,就能形成細胞分裂。這是一個重要的進化事件,因為能夠具有物種特性的生命在本質上必須是一個能自我複製的系統。
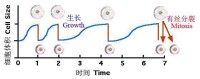
細胞體積隨細胞分裂的變化
基因表達:DNA(複製,周期蛋白,遺傳重組,修復) 翻譯(核糖體亞基,翻譯后修飾,分揀蛋白) DNA/RNA結合蛋白(核糖核蛋白) 結構(1°,2°,2°,結構域,3°,4°) |
| 間期 | G期 S期 G期 |
| M期 | 有絲分裂 早前期 前期 前中期 中期 後期 末期 胞質分裂 |
| 細胞周期檢查點 | 限制點 紡錘體檢查點 複製后檢查點 |
| 其他細胞期 | 凋亡 G期 減數分裂 |
基本信息
- 中文名
- 細胞分裂
- 外文名
- cell division
- 拼音
- xì bāo fēn liè
- 術語類別
- 生物學
- 分類
- 有絲分裂;減數分裂;無絲分裂