生物變異
不遺傳的變異
在豐富多彩的生物界中,蘊含著形形色色的變異現象。在這些變異現象中,有的僅僅是由於環境因素的影響造成的,並沒有引起生物體內的遺傳物質的變化,因而不能夠遺傳下去,屬於不遺傳的變異。有的變異現象是由於生殖細胞內的遺傳物質的改變引起的,因而能夠遺傳給後代,屬於可遺傳的變異。可遺傳的變異有三種來源:基因突變,基因重組,染色體變異。沒有變異就沒有進化,這是從古到今所有進化論者毋庸置疑的共識。但是,關於變異的來源以及如何交織於成種過程(漸變—突變)。
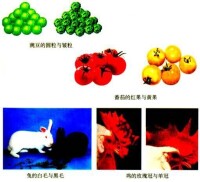
生物變異
人們在對鐮刀型細胞貧血症患者的血紅蛋白分子進行檢查時發現,患者血紅蛋白分子的多肽鏈上,一個谷氨酸被纈氨酸替換。為什麼發生氨基酸分子結構的改變呢?經過研究發現,這是由於控制合成血紅蛋白分子的DNA的鹼基序列發生了改變,這種改變最終導致了鐮刀型細胞貧血症的產生。
基因突變在生物進化中具有重要意義。它是生物變異的根本來源,為生物進化提供了最初的原材料。
引起基因突變的因素很多,可以歸納為三類:一類是物理因素,如X射線、激光等;另一類是化學因素,是指能夠與DNA分子起作用而改變DNA分子性質的物質,如亞硝酸、鹼基類似物等;第三類是生物因素,包括病毒和某些細菌等。
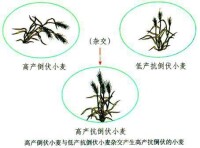
生物變異
無論是低等生物,還是高等的動植物以及人,都可能發生基因突變。基因突變在自然界的特種中廣泛存在。例如,棉花的短果枝,水稻的矮稈、糯性,果蠅的白眼、殘翅,家鴿羽毛的灰紅色,以及人的色盲、糖尿病、白化病等遺傳病,都是突變性狀。自然條件下發生的基因突變叫做自然突變,人為條件下誘發產生的基因突變叫做誘發突變。
它可以發生在生物個體發育的任何時期。一般來說,在生物個體發育的過程中,基因突變發生的時期越遲,生物體表現突變的部分就越少。例如,植物的葉芽如果在發育的早期發生基因突變,那麼由這個葉芽長成的枝條,上面著生的葉、花和果實都有可能與其他枝條不同。如果基因突變發生在花芽分化時,那麼,將來可能只在一朵花或一個花序上表現出變異。
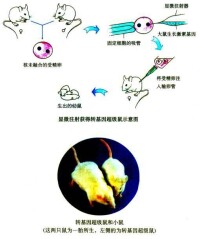
生物變異
第三,在自然狀態下,對一種生物來說,基因突變的頻率是很低的。據估計,在高等生物中,約10五次方到10的八次方個生殖細胞中,才會有1個生殖細胞發生基因突變,突變率是10的負五次方到10的負八次方。
由於任何生物都是長期進化過程的產物,它們與環境條件已經取得了高度的協調。如果發生基因突變,就有可能破壞這種協調關係。因此,基因突變對於生物的生存往往是有害的。例如,絕大多數的人類遺傳病,就是由基因突變造成的,這些病對人類健康構成了嚴重威脅。又如,植物中常見的白化苗,也是基因突變形成的。這種苗由於缺乏葉綠素,不能進行光合作用製造有機物,最終導致死亡。但是,也有少數基因突變是有利的。例如,植物的抗病性突變、耐旱性突變、微生物的抗藥性突變等,都是有利於生物生存的。
一個基因可以向不同的方向發生突變,產生一個以上的等位基因。例如,控制小鼠毛色的灰色基因可以突變成黃色基因,也可以突變成黑色基因。
人工誘變在育種上的應用 人工誘變是指利用物理因素(如X射線、γ射線、紫外線、激光等)或化學因素(如亞硝酸、硫酸二乙酯等)來處理生物,使生物發生基因突變。用這種方法可以提高突變率,創造人類需要的變異類型,從中選擇、培育出優良的生物品種。
基因重組是指在生物體進行有性生殖的過程中,控制不同性狀的基因的重新組合。基因的自由組合定律告訴我們,在生物體通過減數分裂形成配子時,隨著非同源染色體體的自由組合,非等位基因也自由組合,這樣,由雌雄配子結合形成是一種類型的基因重組。在減數分裂形成四分體時,由於同源染色體的非姐妹染色單體之間常常發生局部交換,這些染色體單體上的基因組合,是另一種類型的基因重組。
基因重組是通過有性生殖過程實現的。在有性生殖過程中,由於父本和母本的遺傳特質基礎不同,當二者雜交時,基因重新組合,就能使子代產生變異,通過這種來源產生的變異是非常豐富的。父本與母本自身的雜合性越高,二者的遺傳物質基礎相差越大,基因重組產生變異的可能性也越大。以豌豆為例,當具有10對相對性狀(控制這10對相對性狀的等位基因分別位於10對同源染色體上)的親本進雜交時,如果只考慮基因的自由組合所引起的基因重組,F2可能出現的表現型就有1024種(即2的十次方)。在生物體內,尤其是在高等動植物體內,控制性狀的基因的數目是非常巨大,因此,通過有性生殖產生的雜交後代的表現型種類是很多的。如果把同源染色體的非姐妹染色單體交換引起的基因重組也考慮在內,那麼生物通過有性生殖產生的變異就更多了。
由此可見,通過有性生殖過程實現的基因重組,為生物變異提供了極其豐富的來源。這是形成生物多樣性的重要原因之一,對於生物進化具有十分重要的意義。
基因突變是染色體的某一個位點上基因的改變,這種改變在光學顯微鏡下是看不見的。而染色體變異是可以用顯微鏡直接觀察到的比較明顯的染色體變化,如染色體結構的改變、染色體數目的增減等。
人類的許多遺傳病是由染色體結構改變引起的。例如,貓叫綜合征是人的第5號染色體部分缺失引起的遺傳病,因為患病兒童哭聲輕,音調高,很像貓叫而得名。貓叫綜合征患者的兩眼距離較遠,耳位低下,生長發育遲緩,而且存在嚴重的智力障礙。
在自然條件或人為因素的影響下,染色體發生的結構變異主要有4種:1.染色體中某一片段的缺失;2.染色體增加了某一片段;3.染色體某一片段的位置顛倒了180度;4.染色體的某一片段移接到另一條非同源染色體上。
上述染色體結構的改變,都會使排列在染色體上的基因的數目和排列順序發生改變,從而導致性狀的變異。大多數染色體結構變異對生物體是不利的,有的甚至會導致物體死亡。
一般來說,每一種生物的染色體數目都是穩定的,但是,在某些特定的環境條件下,生物體的染色體數目會發生改變,從而產生可遺傳的變異。染色體數目的變異可以分為兩類:一類是細胞內的個別染色體增加或減少,另一類是細胞內的染色體數目以染色體組的形式成倍地增加或減少。
染色體組 在大多數生物的體細胞中,染色體都是兩兩成對的。例如,果蠅有4對共8條染色體,這4對染色體可以分成兩組,每一組中包括3條常染色體和1條性染色體。就雄果蠅來說,在精子形成的過程中,經過減數分裂,染色體的數目減半,所以雄果蠅的精子中含有一組非同源染色體(Ⅹ、Ⅱ、Ⅲ、Ⅳ 或 Y、Ⅱ、Ⅲ、Ⅳ)
細胞中的一組非同源染色體,它們在形態和功能上各不相同,但是攜帶著控制一種生物生長發育、遺傳和變異的全部信息,這樣的一組染色體,叫做一個染色體組。例如,雄果蠅精子中的這組染色體就組成了一個染色體組。
由受精卵發育而成的個體,體細胞中含有兩個染色體組的叫做二倍體。體細胞中含有三個或三個以上染色體組的叫做多倍體。其中,體細胞中含有三個染色體組的叫做三倍體;體細胞中含有四個染色體組的叫做四倍體。例如,人、果蠅、玉米是二倍體,香蕉是三倍體,馬鈴薯是四倍體。多倍體在植物中很常見,在動物中比較少見。
多倍體產生的主要原因,是體細胞在有絲分裂的過程中,染色體完成了複製,但是細胞受到外界環境條件(如溫度驟變)或生物內部因素的干擾,紡錘體的形成受到破壞,以致染色體不能被拉向兩極,細胞也不能分裂成兩個子細胞,於是就形成染色體數目加倍的細胞。如果這樣的細胞繼續進行正常的有絲分裂,就可以發育成染色體數目加倍的組織或個體。
與二倍體植株相比,多倍體植株的莖稈粗壯,葉片、果實和種子都比較大,糖類和蛋白質等營養物質的含量都有所增加。例如,四倍體葡萄的果實比二倍體品種的大得多,四倍體番茄的維生素C的含量比二倍體的品種幾乎增加了一倍。因此,人們常常採用人工誘導多倍體的方法來獲得多倍體,培育新品種。
人工誘導多倍體的方法很多。目前最常用而且最有效的方法,是用秋水仙素來處理萌發的種子或幼苗。當秋水仙素作用於正在分裂的細胞時,能夠抑制紡錘體形成,導致染色體不分離,從而引起細胞內染色體數目加倍。染色體數目加倍的細胞繼續進行正常的有絲分裂,將來就可以發育成多倍體植株。目前世界各國利用人工誘導多倍體的方法已經培育出不少新品種,如含糖量高的三倍體無子西瓜和甜菜。此外,我國科技工作者還創造出自然界沒有的作物----八倍體小黑麥。
單倍體在生物的體細胞中,染色體的數目不僅可以成倍地增加,還可以成倍地減少。例如,蜜蜂的蜂王和工蜂的體細胞中有32條染色體,而雄蜂的體細胞中只有16條染色體。像蜜蜂的雄蜂這樣,體細胞中含有本物種配子染色體數目的個體,叫做單倍體。
在自然條件下,玉米、高糧、水稻、番茄等高等植物,偶爾也會出現單倍體植株。與正常植株相比,單倍體植株長得弱小,而且高度不育。但是,它們在育種上有特殊的意義。育種工作者常常採用花藥離體培養的方法來獲得單倍體植株,然後經過人工誘導使染色體數目加倍,重新恢復到正常植株的染色體數目。用這種方法得到的植株,不僅能夠正常生殖,而且每對染色體上的成對的基因都是純合的,自交產生的後代不會發生性狀分離。因此,利用單倍體植株培育新品種,只需要兩年時間,就可以得到一個穩定的純系品種。與常規的雜交育種方法相比,明顯縮短了育種年限。
變異從何而來?沒有變異就沒有進化,這是從古到今所有進化論者毋庸置疑的共識。但是,關於變異的來源以及如何交織於成種過程(漸變—突變)卻是歷來進化論者爭論的焦點問題之一。總體來看可分為兩大陣營:以拉馬克和達爾文為代表的博物學家信奉漸變論,他們以“自然無躍進”為箴言,而以德弗里斯和摩爾根為代表的遺傳學家則信奉突變論。
達爾文—環境決定變異
達爾文承認他並不知道變異產生的真正原因,這也是他的進化論常常遭受攻擊的重要原因之一。他在《物種起源》中說,“在較早馴化的動植物中個體之間的差異……一般都比自然狀態下任何一個物種或變種的個體之間的差異更顯著”,他認為將家養動物飼養在不同環境之中就會產生更多的變異,而事實上這可能並不準確,因為,有人甚至認為家畜和栽培植物的品系或品種的個體之間的變異是非常小的。在達爾文的思想中,作為個體變異來源的有性生殖只具有非常次要的作用。
拉馬克則強調動物的主動性適應,他認為生物在趨於完美意志力的驅動下進行變異與適應而獲得遺傳,而這種適應的形式就是所謂的用進廢退。相比之下,達爾文更相信不同的生存環境引起變異(雖然他時而也承認拉馬克的用盡廢退可導致變異),認為這樣的變異主要是隨機的,並通過自然選擇而獲得遺傳。雖然拉馬克強調主動性適應而達爾文更強調被動性適應,但他們都認為生理體制上的變化最終會走向遺傳(這似乎是博物學家的一種直覺)。但是,現代生物學家(特別是分子遺傳學家)則認為這有悖於所謂的中心法則!
魏斯曼—有性生殖是變異的唯一來源
德國遺傳學家魏斯曼(1834年~1914年)認為有性繁殖是能產生無限的個體變異的唯一途徑,而這正是生物種群的主要特徵,在受精時“就好像是兩股遺傳趨勢匯合(結合)在一起。我認為這種結合是可以遺傳的個體性狀的起因,而且我還相信產生這樣的性狀是有性繁殖的真諦。這一過程(有性繁殖)的目的是產生作為自然選擇形成新物種的素材的那個個體差異”(邁爾1990)。魏斯曼將有性繁殖作為個體變異的唯一途徑未免過於誇張(雖然是主要途徑),但他意識到有性生殖對個體變異的重要性還是值得肯定的,雖然那時還不可能認識到有性生殖的分子機制和本質。謝平(2013)指出,有性生殖也不是如魏斯曼所說的是為了產生變異:進行有性生殖的物種是以群體基因庫的形式保存著變異性狀(如通過等位基因),此外,雖然有性生殖在減數分裂中容易導致基因變異,但它只是一種偶然的結果而非原因。
柏格森—生命的原始衝動是變異的根本原因
法國哲學家柏格森(1907)認為,“……生命的原始衝動(original impetus),通過連接代際間隔的成熟有機體從前一代胚芽傳給下一代胚芽。這種衝動沿著進化的路線持續,被這些路線分開,它就是變異的根本原因,至少是那些被有規則地傳遞的變異的根本原因,是那些積累和創造新物種的變異的根本原因。總之,從物種開始從共同的祖先分化起,它們就在各自的進化過程中強化了自己的差異。然而,在某些確定的點上,它們又可能出現共同的演變;實際上,倘若接受‘共同衝動’的假定,它們就必須如此”。
那什麼是這個原始衝動呢?柏格森說,“所謂原始衝動,就是一種使生命得以發展的內在衝動,其形式越來越複雜,其最終目標越來越高”。這不就是拉馬克所說的一切生命就具有一種內稟的趨於向高級生命進化的趨勢么?
薛定諤—只有突變才能遺傳
奧地利理論物理學家薛定諤在其1944年出版的《生命是什麼》一書中,對達爾文的“變異”進行了猛烈的抨擊,他說,“今天我們已經明確地知道,達爾文是錯誤地把即使在最純的群體里也會出現的細微的、連續的、偶然的變異,當作是自然選擇的原始材料。因為後來已經證明,這些變異是不遺傳的”(Schrödinger 1944),另一方面,他對德弗里斯的突變論大加讚賞,甚至將其吹捧為“生物學的量子論”!薛定諤徹底否定達爾文的連續變異理論,認為那些細微的、連續的的變異是不可能遺傳的。
但薛定諤也說,“突變肯定是遺傳寶庫中的一種變化……突變是達爾文描述的自然選擇的合適的原料,在此過程中,通過不適者被淘汰、最適者生存從而產生新種……只要用‘突變’來代替‘細微的偶然變異’(正如在量子論中用‘量子躍遷’來代替‘能量的連續轉移’),達爾文學說的其他方面是不需要作什麼修改的”(Schrödinger 1944)。筆者認為,物理學家的量子理論看上去與分子遺傳學家的基因突變理論十分情投意合,這在較短的時間尺度上似乎也無可非議,但是,這在本質上也是一種唯基因論(基因萬能、基因決定論)!
當然,薛定諤也意識到,“如果突變是如此的頻繁,以致有很多的機會,比如說,在同一個體內出現了十幾個不同的突變,而其中有害的突變又總比有利突變佔優勢,那麼,物種非但不會通過選擇而得到改良,反而會停滯,甚至會消亡。由基因的高度持久性形成的相當程度的保守性是十分必要的”(Schrödinger 1944)。
事實上,生物體內的生化機製表明,物種並未放任變異,它們時刻都在努力減少DNA複製過程中的錯誤以維護遺傳的穩定性,這是因為生物界普遍存在著DNA的修復機制。但是DNA修復又需付出能量與時間的代價,因此是對少量錯誤的一種“無奈”的放棄(正是這種“無奈”,也是生物界得以如此繁盛的重要契機之一)。此外,有時不能保證對一切錯誤的完全修復,譬如,當在DNA雙鏈的同樣位點同時出錯(無論是複製錯誤還是受到物理化學條件的誘導)時,則修復就不再可能,要麼由於它是有害的突變而被淘汰,或中性的突變而被保留,或有利的突變而被保留,如果足夠幸運的話,甚至可在種群中擴散。或許可以設想,如果再多一份拷貝(二倍體),或許就能保證更好的遺傳穩定性,但事實上,這樣的結果往往會導致一個新物種的出現!
基本信息
- 中文名
- 生物變異
- 外文名
- biotic variation
- 特點
- 普偏性,隨機性,多害性
- 類型
- 專業用語
- 來源
- 基因突變,基因重組等
- 遺傳的變異
- 3種
- 變異現象
- 形形色色
- 範圍
- 生物
- 造成
- 環境因素
- 變異類型
- 結構變異,數目變異
- 屬於
- 不遺傳的變異