甲殼亞門
甲殼綱門
甲殼亞門是節肢動物門中的一個亞門,由於亞門下物種用鰓呼吸,過去曾與三葉蟲綱一起列為有鰓亞門(Branchiata),後來又被列為有顎亞門(Mandibulata),後有更多學者主張應獨立成甲殼亞門(Crustacea)。甲殼類動物約有31000多種,包括我們熟悉的水蚤、劍水蚤、豐年蟲、對蝦、螯蝦、龍蝦、蟹等。
目錄
| 海洋或淡水生活,極少數在潮濕的陸地上生活 |
| 身體分為頭胸部和腹部 |
| 體節及附肢較多,附肢大多保持雙枝型,有2對觸角 |
| 消化系統常為一根直管,胃內具有幾丁質構造的胃磨 |
| 呼吸器官為鰓 |
| 排泄器官為觸角腺(成體)、小顎腺(幼體) |
| 低等種類神經系統保持梯形,高等種類為鏈狀 |
| 一般為雌雄異體,發育經變態(無節幼體等) |
甲殼亞門在形態結構上表現出多樣性,例如體形大小、體節數目、體軀劃分及附肢的分化都有較大的差異。在生活方式上絕大多數為海洋生,為海洋浮遊動物的重要成員,常被譽為海洋中的昆蟲,少數侵入淡水。是構成水生食物鏈中的重要環節。
也有極少數進入陸地生活,但不能脫離潮濕環境。
甲殼類動物都具有兩對觸角,這一點區別於其它節肢動物(有螯肢亞門無觸角,三葉蟲亞門及單肢亞門僅有一對觸角,相當於甲殼類的小觸角)。甲殼類動物的附肢基本上都是雙肢型,但在不同的體區有不同的形態與機能的分化,其中至少有三對口后的附肢作為顎。
甲殼類動物生活史中由卵孵出後為無節幼蟲(nauplius),僅有三個體節,三對附肢,相當於成體的前三對附肢,即兩對觸角及一對大顎。經多次蛻皮不斷地在肛門前部增加體節數,原始的種類體節可多達數十節,高等的種類體節數減少並固定,體區劃分清楚。
此外有些種類頭部和胸部可能也會融合到一起,形成頭胸部。
所有的甲殼亞門的幹細胞分裂時的特徵相同。
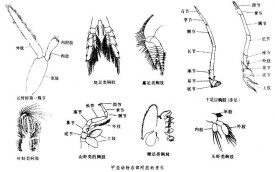
外部形態 甲殼動物的形態有著較多的變化,隨不同的種類其體節、分區都有很大的差異。低等的甲殼類一般體型小,只有幾mm到2~3cm,體節數多且不固定,但分節明顯,各節形態相似,體表外骨骼薄而透明。例如豐年蟲(Chirocephalus),身體分為頭部與軀幹部,沒有背甲。又如鱟蟲(Apus),體節數可多達50多節,胸、腹部分界不明顯,有背甲(carapace),其頭部體節與胸部體節癒合成頭胸部(cephalothorax)。還有的種類體表完全被石灰質骨板包圍,並營固著生活,例如藤壺(Balanus)。低等甲殼類生物形態的多樣性在細分的綱中會有較為詳細的介紹。高等的甲殼類動物,即軟甲綱動物身體的節數固定,分區明顯,一般是由20~21節組成。例如蝦的頭部5節,胸部8節,腹部6節及1尾節。部分胸部體節與頭部合併,形成頭胸部,之間分界消失。頭胸部背面具有一發達的背甲,也稱頭胸甲,蓋在身體的背部,或向兩側延伸也蓋住體側的部分附肢及鰓,因此也稱為鰓蓋,其下的空腔稱鰓腔。頭胸甲是由頭及胸部的體褶共同分泌的外骨骼所形成。甲殼類動物有發達的附肢,除了觸角與尾節之外,每個體節都有一對雙肢型附肢。隨著不同種及不同的著生部位,附肢有多種形態及功能的分化。原始的種類附肢數目隨體節數也較多,但分化很少,多呈葉片狀(foliaceous),它相似於環節動物的疣足,與體軀無關節而是直接相連,可分為內葉(內肢)(endopodite)與外葉(外肢)(exopodite),基部的突起形成了副葉(epipodite),具鰓的功能。豐年蟲、鱟蟲都是這種原始的葉片狀附肢。高等的甲殼類動物附肢隨體節數減少而減少,均與軀幹部以關節相連,附肢本身亦分節。與軀幹相關節的部分稱原肢節(protopodite),由原肢節再生出內肢與外肢,蝦、蟹腹部的游泳足便是這樣。多數附肢、特別是胸部附肢原肢節可分為兩節,與體軀相關節的稱為底節(coxopodite),其下一節為基節(basipodite)。由基節再分出內肢與外肢。內肢可再分成5節,由基部到端部依次為座節(ischiopodite)、長節(meropodite)、腕節(carpopodite),掌節(propodite)及指節(dactylopodite)。原肢節上可長出旁枝,稱副肢。在原肢節的內外邊緣還可長出突起,稱外小葉(exite)及內小葉(endite),一些種類外小葉變成了鰓,靠近口部附肢的內小葉變成了顎。如果外肢節減小或退化,則變成單肢型附肢。以沼蝦(Macrobrachium)為例,共有19對附肢,它們的分佈是:頭部5對,其中第一對為小觸角(antenn-ules),單肢型,同源於其他亞門的觸角,第二對為大觸角(antennae),均具感覺功能。第三對為大顎(mandibles),其原肢節大而堅硬,內面具齒,用以咀嚼。后二對為第一及第二小顎(maxillae),呈葉片狀,用以把持食物,第二小顎的外肢擴大形成顎基葉(gnathobasic lobes),用以激動水在鰓腔中的流動。后三對附肢位於口后。口的前、後有體壁形成的突起,構成上唇(labrum)及下唇(labium)。胸部8對附肢,前三對形成顎足(maxillipeds),雙肢型,具副肢及鰓,用以呼吸、感覺及把持食物。胸部的后5對為步行足(pereiopoda),外肢退化或消失,原肢2節,內肢5節,末端具螯或爪,具步行、取食、呼吸及防衛的功能。腹部6對附肢,原始的雙肢型,原肢節及內肢節均不分節,用以游泳,稱腹足(pleopoda)。雌性的可用以攜卵,雄性的第1~2對附肢用以輸送精子。第6對附肢與尾節合併成尾扇,具舵的功能。
甲殼動物的形態有著較多的變化,隨不同的種類其體節、分區都有很大的差異。低等的甲殼類一般體型小,只有幾mm到2~3cm,體節數多且不固定,但分節明顯,各節形態相似,體表外骨骼薄而透明。例如豐年蟲(Chirocephalus),身體分為頭部與軀幹部,沒有背甲。又如鱟蟲(Apus),體節數可多達50多節,胸、腹部分界不明顯,有背甲(carapace),其頭部體節與胸部體節癒合成頭胸部(cephalothorax)。還有的種類體表完全被石灰質骨板包圍,並營固著生活,例如藤壺(Balanus)。低等甲殼類生物形態的多樣性在細分的綱中會有較為詳細的介紹。高等的甲殼類動物,即軟甲綱動物身體的節數固定,分區明顯,一般是由20~21節組成。例如蝦的頭部5節,胸部8節,腹部6節及1尾節。部分胸部體節與頭部合併,形成頭胸部,之間分界消失。頭胸部背面具有一發達的背甲,也稱頭胸甲,蓋在身體的背部,或向兩側延伸也蓋住體側的部分附肢及鰓,因此也稱為鰓蓋,其下的空腔稱鰓腔。頭胸甲是由頭及胸部的體褶共同分泌的外骨骼所形成。甲殼類動物有發達的附肢,除了觸角與尾節之外,每個體節都有一對雙肢型附肢。隨著不同種及不同的著生部位,附肢有多種形態及功能的分化。原始的種類附肢數目隨體節數也較多,但分化很少,多呈葉片狀(foliaceous),它相似於環節動物的疣足,與體軀無關節而是直接相連,可分為內葉(內肢)(endopodite)與外葉(外肢)(exopodite),基部的突起形成了副葉(epipodite),具鰓的功能。豐年蟲、鱟蟲都是這種原始的葉片狀附肢。高等的甲殼類動物附肢隨體節數減少而減少,均與軀幹部以關節相連,附肢本身亦分節。與軀幹相關節的部分稱原肢節(protopodite),由原肢節再生出內肢與外肢,蝦、蟹腹部的游泳足便是這樣。多數附肢、特別是胸部附肢原肢節可分為兩節,與體軀相關節的稱為底節(coxopodite),其下一節為基節(basipodite)。由基節再分出內肢與外肢。內肢可再分成5節,由基部到端部依次為座節(ischiopodite)、長節(meropodite)、腕節(carpopodite),掌節(propodite)及指節(dactylopodite)。原肢節上可長出旁枝,稱副肢。在原肢節的內外邊緣還可長出突起,稱外小葉(exite)及內小葉(endite),一些種類外小葉變成了鰓,靠近口部附肢的內小葉變成了顎。如果外肢節減小或退化,則變成單肢型附肢。以沼蝦(Macrobrachium)為例,共有19對附肢,它們的分佈是:頭部5對,其中第一對為小觸角(antenn-ules),單肢型,同源於其他亞門的觸角,第二對為大觸角(antennae),均具感覺功能。第三對為大顎(mandibles),其原肢節大而堅硬,內面具齒,用以咀嚼。后二對為第一及第二小顎(maxillae),呈葉片狀,用以把持食物,第二小顎的外肢擴大形成顎基葉(gnathobasic lobes),用以激動水在鰓腔中的流動。后三對附肢位於口后。口的前、後有體壁形成的突起,構成上唇(labrum)及下唇(labium)。胸部8對附肢,前三對形成顎足(maxillipeds),雙肢型,具副肢及鰓,用以呼吸、感覺及把持食物。胸部的后5對為步行足(pereiopoda),外肢退化或消失,原肢2節,內肢5節,末端具螯或爪,具步行、取食、呼吸及防衛的功能。腹部6對附肢,原始的雙肢型,原肢節及內肢節均不分節,用以游泳,稱腹足(pleopoda)。雌性的可用以攜卵,雄性的第1~2對附肢用以輸送精子。第6對附肢與尾節合併成尾扇,具舵的功能。
甲殼綱
體壁與色素 體壁結構與其它節肢動物相似,只是某些甲殼類如蝦、蟹的外骨骼有相當程度的鈣化,主要是因為上表皮及外表皮均有相當多鈣鹽的沉積。另外在表皮細胞的下面有皮下腺(tegumentalgland)和色素細胞(ehromatophores)。皮下腺是一群具分泌能力的腺細胞,並有長管穿過外骨骼開口在上表皮的表面,它的功能還不十分清楚。色素細胞是位於表皮細胞下面的結締組織中的一些呈放射狀分支的細胞,放射狀的突起不能伸縮,但細胞中含有大量的色素顆粒,呈白、紅、黃、藍、褐、黑等色。根據生理狀況的不同,顆粒或分散在整個細胞,或集中在細胞中心。其中紅、黃、藍色是類胡蘿蔔素,來自於食物,在生活狀態時,這些色素常與蛋白質呈結合狀態而表現出不同的顏色。當蝦、蟹加熱煮熟時,體表出現美麗的紅色,這是因為加熱使蛋白色沉澱出來而留下紅色化合物——蝦青素(astaxanthin)的緣故。事實上,一個單個的色素細胞可以包括一種到幾種不同顏色的色素,任何一種色素都可獨立移動。多色素的色素細胞僅出現在蝦中。體色的改變有兩種形式:一種是形態變色(morphologicalcolorchange),它涉及色素細胞中色素的丟失或形成,或是由於動物長期在一固定的環境及光照條件下細胞數目的改變而形成;另一種形式是生理變色(physiological color change)它是指對環境的迅速的顏色適應,來自於色素細胞中色素顆粒的分散與集中。最普通的生理變色是體色深淺的改變,這在許多蟹中可以發現。而蝦類常有更廣泛的顏色變化,例如一種小長臂蝦(Palaemonetes),色素細胞中含有紅、黃、藍三種色素顆粒,通過其中任一色素顆粒的獨立移動使之可以適應任何顏色的背景。許多蝦類都具有這種能力。色素顆粒在色素細胞中的移動受眼柄中分泌的激素(hormones)控制。例如許多蝦的色素細胞含有紅、黃、藍和白色素,移走眼柄,紅、黃色素擴散而使體色變暗。如注射眼柄的激素提取物,則白色素擴散而使體色迅速變淺。因此,對每種色素可能都存在著一對對抗性的促色素細胞素(chromatophorotrophins),其中一個處於分散狀態,另一個處於集中狀態。體色深淺的改變,取決於一對對抗性促色素細胞素中的哪一個處於擴散狀態以及哪一個處於集中狀態。
體壁結構與其它節肢動物相似,只是某些甲殼類如蝦、蟹的外骨骼有相當程度的鈣化,主要是因為上表皮及外表皮均有相當多鈣鹽的沉積。另外在表皮細胞的下面有皮下腺(tegumentalgland)和色素細胞(ehromatophores)。皮下腺是一群具分泌能力的腺細胞,並有長管穿過外骨骼開口在上表皮的表面,它的功能還不十分清楚。色素細胞是位於表皮細胞下面的結締組織中的一些呈放射狀分支的細胞,放射狀的突起不能伸縮,但細胞中含有大量的色素顆粒,呈白、紅、黃、藍、褐、黑等色。根據生理狀況的不同,顆粒或分散在整個細胞,或集中在細胞中心。其中紅、黃、藍色是類胡蘿蔔素,來自於食物,在生活狀態時,這些色素常與蛋白質呈結合狀態而表現出不同的顏色。當蝦、蟹加熱煮熟時,體表出現美麗的紅色,這是因為加熱使蛋白色沉澱出來而留下紅色化合物——蝦青素(astaxanthin)的緣故。事實上,一個單個的色素細胞可以包括一種到幾種不同顏色的色素,任何一種色素都可獨立移動。多色素的色素細胞僅出現在蝦中。體色的改變有兩種形式:一種是形態變色(morphologicalcolorchange),它涉及色素細胞中色素的丟失或形成,或是由於動物長期在一固定的環境及光照條件下細胞數目的改變而形成;另一種形式是生理變色(physiological color change)它是指對環境的迅速的顏色適應,來自於色素細胞中色素顆粒的分散與集中。最普通的生理變色是體色深淺的改變,這在許多蟹中可以發現。而蝦類常有更廣泛的顏色變化,例如一種小長臂蝦(Palaemonetes),色素細胞中含有紅、黃、藍三種色素顆粒,通過其中任一色素顆粒的獨立移動使之可以適應任何顏色的背景。許多蝦類都具有這種能力。色素顆粒在色素細胞中的移動受眼柄中分泌的激素(hormones)控制。例如許多蝦的色素細胞含有紅、黃、藍和白色素,移走眼柄,紅、黃色素擴散而使體色變暗。如注射眼柄的激素提取物,則白色素擴散而使體色迅速變淺。因此,對每種色素可能都存在著一對對抗性的促色素細胞素(chromatophorotrophins),其中一個處於分散狀態,另一個處於集中狀態。體色深淺的改變,取決於一對對抗性促色素細胞素中的哪一個處於擴散狀態以及哪一個處於集中狀態。
當蝦、蟹煮熟時,只留下蝦青素。
運動與取食甲殼類動物的運動方式與生活方式密切相關,小型的甲殼類動物保留了祖先的水底表面懸浮取食的習性,因此它們以某些附肢的打動以推動身體前進。例如水蚤(Daphnia)、劍水蚤(Cyclops)等以第二對觸角像槳一樣在水中划動,推動身體呈跳躍式的前進。在水中游泳生活的種類,其游泳肢具有剛毛繸,以增加抗水表面。大型游泳生活的甲殼類動物腹部具有發達的肌肉,靠肌肉的收縮推動身體前進,例如各種蝦。爬行生活的蟹類,雖然某些種還保留了游泳的能力,但更多的附肢已變得適合於爬行及挖掘。甲殼類動物的食物類型及取食習性也表現出多樣性,有植食的、肉食的、腐食的以及寄生生活的,取食方式及相應的附肢因種而有很大不同。許多小型的甲殼類動物都是過濾取食,它們以浮游生物或沉渣為食,這種取食方式可能與運動相關。其頭部及軀幹前端的附肢布滿細微的剛毛,不僅用以激動水流,同時也作為過濾器以收集水中的食物顆粒。剛毛間空間的大小決定了食物顆粒的大小。剛毛收集的食物顆粒經附肢腹面的食物溝送入口。大型的甲殼類動物多為捕食性取食,前端的步足適合於捕獲及撕裂食物,大小顎及顎足用以咬切及把持食物。大小顎及上、下唇組成口器。十足目的第三對顎足排列在口外組成門狀的保護器。消化道呈直管狀,口位於前端腹面,前腸包括口、食道、膨大的噴門胃(cardiac stomach)及幽門胃(pyloric stomach)。在噴門胃及幽門胃的胃壁上有幾丁質硬化形成的嵴及獨立的齒,用以研磨食物,並有肌肉控制胃壁及齒的運動。取食微小食物的種類嵴與齒不發達。幽門胃的胃壁多褶皺而使胃腔減小,褶皺上生有大量的剛毛而形成過濾系統,使小的食物顆粒進入消化盲囊,大的顆粒進入中腸。幽門胃後為中腸,中腸的前端有一對或數對消化盲囊。在十足目一對消化盲囊變成實體的消化腺,稱為肝胰腺(hepatopancreas),它幾乎充滿了頭胸部,它能產生消化酶並進行食物的消化,也是糖元、脂肪及鈣的吸收與貯存的場所。中腸也是消化及吸收的場所。中腸后經后腸以肛門開口在尾節腹面。
呼吸小型甲殼類動物沒有專門的呼吸器官,而是用體表進行氣體交換。某些種雖有附肢特化成鰓,但仍以體表呼吸為主,例如鰓足類。軟甲綱都以鰓作為呼吸器官。典型的鰓與胸部體節的側壁及附肢相關,鰓的數目、結構、位置因種而不同。原始的胸部每個體節應有4對鰓,其中一對附著在體側壁上,稱側鰓(pleurobranch),2對固著在附肢底節與體壁相連的關節膜上,稱關節鰓(arthrobranch),一對附著在附肢基節上,稱足鰓(podobranch)。但現存種類實際上都少於32對鰓,例如深對蝦(Benthesicymus)只有24對,螯蝦只有17對,海產蟹類一般只有9對,最少的豆蟹(Pinnotheres)只有3對。所有的鰓均位於鰓室中,並被頭胸甲延伸形成的鰓蓋(branchiostegite)所遮蓋。生活時靠附肢、特別是第2顎足的顎基葉的打動造成水流。水可由鰓蓋的腹緣及后緣流入鰓室,或由附肢基部及鰓蓋后緣流入。在蟹類,僅由螯肢的基部流入。水都由頭胸甲前端兩側流出。水在鰓腔中的循環時,使鰓得以進行氣體交換。蟹類三對顎足副肢及部分步足的基節上具有大量的剛毛,用以清潔鰓及過濾水中的雜質,以防鰓被沉渣污染。鰓的結構可分為三類:支鰓(dendrobranch),由鰓軸向兩側伸出側支,或側支再分支,例如對蝦類;毛鰓(trichobranch),圍繞鰓軸具毛狀或絲狀排列,例如大多數的長尾蝦及螯蝦等;葉鰓(phyllobranch),沿鰓軸向兩側伸出葉片狀鰓頁,例如長臂蝦(palaemon)、寄居蟹類及蟹類,鰓軸中有入鰓血竇及出鰓血竇,流經鰓絲或鰓葉時進行氣體的交換。陸生及半陸生的種類仍然保留了較大的氣體交換的表面積,例如海岸邊生活的招潮蟹(Uca);它攜帶水分在鰓腔中,並在第3~4對足上有呼吸孔,以允許氣體進入,氣體的交換仍在鰓腔中進行。真正陸生的種類如椰子蟹(Birgus)、鼠婦(Porcellio)等,其鰓腔壁變成囊室而形成肺。
循環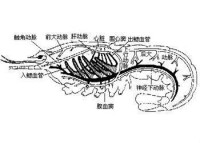 切甲類一般沒有循環的結構,血液在血腔中通過身體的運動而循環。軟甲類都有循環系統,也屬開放式,包括心臟、血管及血竇。心室的形態因種而異,或呈囊狀、管狀,或呈實體狀,均位於胸部背面的圍心竇中。心室上有心孔,血液由心孔進入心室。低等的種類,血液由心室出來,經短而不分支的動脈進入組織,再經靜脈血竇進入鰓,行氣體交換后再回到圍心竇,最後回到心室。十足目的心室呈實體狀,有三對心孔。蝦的心孔在心室的前側角、后側角各一對,背面一對。由心室向前發出五條動脈;一條眼動脈(ophthalmic artery);一對觸角動脈(antennary arteries)及一對肝動脈(hepatic arteries)。由心臟向後發出一條腹動脈(abdominalartery),沿背中線後行。此外,還有一條胸直動脈(sternal artery),由心臟下面發出,沿消化道側面直達腹面穿過神經索分成兩支,稱為神經下動脈(subneural artery),一支向前,一支向後。這些血管再分支,進入組織,然後流入胸竇,進入鰓後行氣體交換,最後再流回圍心竇,經心孔再流回心室。甲殼類的血漿中溶解有血青蛋白,故血液多呈淡藍色。血液中含有透明的及大的顆粒狀的變形細胞,其數目因種、年齡及生理狀態而不同,但一般在數百—數千/cm3血液中。血細胞不僅具吞噬能力,也參與血液的凝結。在刺激條件下,如斷肢時某些變形細胞可以分解、破裂,並釋放出某些物質將血漿中的纖維蛋白原(fibrinogen)轉變成纖維蛋白(fibrin),結果出現凝結的血漿島並被沉澱,形成血塊。一些小型的甲殼類在血液及組織液中含有少量血紅蛋白,它與氧的親和力較大,適合於在低氧條件下呼吸。
切甲類一般沒有循環的結構,血液在血腔中通過身體的運動而循環。軟甲類都有循環系統,也屬開放式,包括心臟、血管及血竇。心室的形態因種而異,或呈囊狀、管狀,或呈實體狀,均位於胸部背面的圍心竇中。心室上有心孔,血液由心孔進入心室。低等的種類,血液由心室出來,經短而不分支的動脈進入組織,再經靜脈血竇進入鰓,行氣體交換后再回到圍心竇,最後回到心室。十足目的心室呈實體狀,有三對心孔。蝦的心孔在心室的前側角、后側角各一對,背面一對。由心室向前發出五條動脈;一條眼動脈(ophthalmic artery);一對觸角動脈(antennary arteries)及一對肝動脈(hepatic arteries)。由心臟向後發出一條腹動脈(abdominalartery),沿背中線後行。此外,還有一條胸直動脈(sternal artery),由心臟下面發出,沿消化道側面直達腹面穿過神經索分成兩支,稱為神經下動脈(subneural artery),一支向前,一支向後。這些血管再分支,進入組織,然後流入胸竇,進入鰓後行氣體交換,最後再流回圍心竇,經心孔再流回心室。甲殼類的血漿中溶解有血青蛋白,故血液多呈淡藍色。血液中含有透明的及大的顆粒狀的變形細胞,其數目因種、年齡及生理狀態而不同,但一般在數百—數千/cm3血液中。血細胞不僅具吞噬能力,也參與血液的凝結。在刺激條件下,如斷肢時某些變形細胞可以分解、破裂,並釋放出某些物質將血漿中的纖維蛋白原(fibrinogen)轉變成纖維蛋白(fibrin),結果出現凝結的血漿島並被沉澱,形成血塊。一些小型的甲殼類在血液及組織液中含有少量血紅蛋白,它與氧的親和力較大,適合於在低氧條件下呼吸。
軟甲類都有循環系統,也屬開放式
排泄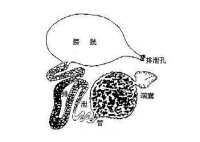 甲殼類的排泄器官是一對觸角腺(antennal glands),或是一對小顎腺(maxillary glands),因位於第二觸角或小顎基部而得名。幼蟲期二種腺體常同時存在,成蟲期往往只有其中的一種。也有兩種同時存在的種類,如某些介形類。其結構相似於有螯類的基節腺,也是由殘留的按節排列的后腎形成。由基部的囊及排泄管組成。在軟甲類,觸角腺或小顎腺有了高度的發展,端囊包括一個小囊(它從周圍的血液中收集含廢物的液體成分)及一個大的腺體部分,它是囊壁褶皺形成的海綿狀組織,是重要的重吸收部位。由腺體部再通出排泄管,其後端膨大形成了膀胱,最後經一短管在第二觸角的基部開口到體外。甲殼類的代謝產物主要是氨及少量的尿酸。雖然觸角腺稱為排泄器官,但大量的含氮廢物是通過鰓或體壁的擴散作用而排出體外。另外,在鰓軸處及足的基部有一些腎細胞(nephrocytes),這些細胞可以攝取及貯存尿酸形成顆粒,成為色素細胞的一部分,或沉澱在體壁中,隨蛻皮時排出。而觸角腺的主要機能在於維持及調節體內離子濃度及液體壓力的平衡。當體內血液中水分含量增高時,觸角腺的過濾作用也增強,並形成較多的尿而排出體外。例如一種蟹Carcinus,如果生活在海水中(含鹽量34‰),每日產生相當於3.6%體重的尿,如果生活在鹹水中(14‰),則每日產生相當於1/3體重的尿。這說明觸角膜的機能在於維持穩定的體積常數,而不管外界環境中的鹽度含量。大多數十足目所產生的尿與血液是等滲的,而且是能調節鹽度的,因為它們的鰓可以由水流中攝取離子以補償由尿中失去的離子。另外,排泄管也具有重吸收作用,以產生低滲的尿排出體外。
甲殼類的排泄器官是一對觸角腺(antennal glands),或是一對小顎腺(maxillary glands),因位於第二觸角或小顎基部而得名。幼蟲期二種腺體常同時存在,成蟲期往往只有其中的一種。也有兩種同時存在的種類,如某些介形類。其結構相似於有螯類的基節腺,也是由殘留的按節排列的后腎形成。由基部的囊及排泄管組成。在軟甲類,觸角腺或小顎腺有了高度的發展,端囊包括一個小囊(它從周圍的血液中收集含廢物的液體成分)及一個大的腺體部分,它是囊壁褶皺形成的海綿狀組織,是重要的重吸收部位。由腺體部再通出排泄管,其後端膨大形成了膀胱,最後經一短管在第二觸角的基部開口到體外。甲殼類的代謝產物主要是氨及少量的尿酸。雖然觸角腺稱為排泄器官,但大量的含氮廢物是通過鰓或體壁的擴散作用而排出體外。另外,在鰓軸處及足的基部有一些腎細胞(nephrocytes),這些細胞可以攝取及貯存尿酸形成顆粒,成為色素細胞的一部分,或沉澱在體壁中,隨蛻皮時排出。而觸角腺的主要機能在於維持及調節體內離子濃度及液體壓力的平衡。當體內血液中水分含量增高時,觸角腺的過濾作用也增強,並形成較多的尿而排出體外。例如一種蟹Carcinus,如果生活在海水中(含鹽量34‰),每日產生相當於3.6%體重的尿,如果生活在鹹水中(14‰),則每日產生相當於1/3體重的尿。這說明觸角膜的機能在於維持穩定的體積常數,而不管外界環境中的鹽度含量。大多數十足目所產生的尿與血液是等滲的,而且是能調節鹽度的,因為它們的鰓可以由水流中攝取離子以補償由尿中失去的離子。另外,排泄管也具有重吸收作用,以產生低滲的尿排出體外。
甲殼類的排泄器官是一對觸角腺,或是小顎腺
神經與感官甲殼類的神經系統也是鏈狀神經。原始的種類腦後的神經索呈雙鏈狀,例如某些鰓足類,即在每一個體節各有一對神經節,彼此分離,之間有成雙的橫神經相連,前後神經節各自成鏈,因此形成雙鏈。但大多數甲殼類不存在這種原始的結構,而是雙鏈癒合成單鏈。如各種蝦、蟹類胸腹部神經節常與食道下神經節合併成團塊狀,腦神經節也更集中。由腦發出神經到觸角、眼,圍咽神經環等。腹神經索中的每個神經節分出三對側神經,其中前兩對是運動與感覺神經,支配附肢與體節,最後一對是運動神經,支配體壁的肌肉。許多甲殼類的神經索中也有巨大神經纖維,能快速傳導衝動產生反應。其纖維的直徑可達200μm。一般在神經索的背中、背側各有兩個巨大纖維,前者是由腦神經細胞發出,其纖維達神經索的全長,後者是由神經節的細胞發出,所以刺激任何一種巨大纖維都可引起身體的快速運動。蝦正是以這種機制完成快速跑動、取食、繁殖、自切等各種反射活動的。甲殼類具兩種類型的眼,即中眼(median eyes)與複眼(compound eyes)。中眼一個,是無節幼蟲期所普遍有的,成體后或者存在,或者消失。在低等的種類成為成蟲期僅有的眼,如橈足類。它由3~4個具色素杯的小眼組成,小眼由少量的視覺細胞組成,一般沒有晶體狀結構。中眼的機能可能僅在於測知光源,使動物趨向於水的表層或低層。大多數成體具一對複眼,分別位於頭的兩側。多數種類眼位於眼柄(peduncle)上,眼柄由2~3節組成,一般可以活動。眼的表面角膜有很大的凸度,其弧面可達180°,再加上眼柄的運動,因此具有很大的視覺範圍。複眼中的小眼數少的僅有20多個,多的可達15000個,如螯龍蝦(Homarus)。大多數甲殼類的複眼對強、弱光均有色素屏幕效應,也就是說,生活在光照條件下的種類(陸地、淺水)一般具並列眼,而一些洞穴及深水生活的種類由於色素屏幕不發達而具重疊眼。另外,從眼的結構研究表明,甲殼類的複眼能夠識別物體的形狀、大小及顏色變化。例如寄居的蟹類可以區別不同顏色的螺殼,一些蝦可以在紅、黃、褐色的背景中生存,而不適合深色環境。十足目常有一對平衡囊,位於觸角的基部,或足的基部或尾節,它由外胚層凹陷形成,內有平衡石,外有開口。平衡石或由外界攝入,或由細胞分泌。平衡囊也隨蛻皮而脫落,蛻皮后再形成。囊的底部內壁有感覺毛,或與平衡石直接接觸,或通過囊內的液體而感受刺激。平衡囊是其重力感受器,並與運動相關,以調節身體平衡。除了以上結構,甲殼動物外骨骼的表面分佈有各種感覺毛,有多種機能。最常見的是分佈於附肢上的觸覺感受器,用以測知水流及其方向,它與外界環境直接接觸。還有存在於觸角及圍繞口部附肢上的化學感受器,特別多地分佈在小觸角的外鞭上,由觸角上的雙極神經細胞所支配。分佈於觸角的還有溫度感受器。在腹部肌肉兩側,還有一些由肌肉細胞改變形成的本體感受器(proprioceptors),它們的機能還不十分清楚。
生殖與發育大多數的甲殼動物是雌雄異體,但也有一些種類為雌雄同體,例如蔓足類、寄生的等足類等。還有一些種類為孤雌生殖,例如鰓足類、介形類等。甲殼動物的生殖腺(精巢、卵巢)通常是一對長形器官,位於胸部或腹部背面,或同時存在於胸腹部。精巢一對,背部靠近或癒合成一整塊。一對輸精管。有的種類以精莢進行傳遞精子,這時其輸精管的後端部分變成腺狀,以便於精莢的形成。在十足類輸精管末端形成肌肉質的射精管(ejaculatoryduct),最後以生殖孔開口在最後一對胸部附肢的基部,或最後胸節的腹板上。十足目雄性的第1~2對腹足特化,用以傳導精子。大多數甲殼類的精子是無尾的,呈球形或星形等非典型精子形態。卵巢的結構與位置相似於精巢。一對輸卵管。十足目的輸卵管末端聯合,還形成受精囊及陰道,最後以雌性生殖孔開口在附肢基部。受精囊開口到外界,並在最後1~2對胸節腹板突起形成一盤狀。在十足目中出現某些種的性二型現象。例如招潮蟹,其雄性的一個附肢的爪特別發達,而雌性具二爪且相等大小。有的種雌雄個體大小相異。交配前也常有性引誘(sexual attraction)及追逐行為(courtship)。引誘行為或是視覺的,或是聽覺的。例如,招潮蟹在交配之前雄性在雌性面前不斷的搖動,其搖動的方式在同一種是固定的。另外,雄性的爪變成紅色,以便於雌性識別。它還以爪敲打胸腹板,或敲打步足以產生聲音信號,其敲打次數及間隔長短也因種而異。這種敲打聲在水中可傳到50~100cm之外,而被同種雌性個體足部本體感受器所接受,以吸引雌性到雄性的穴中進行交配,並在此產卵孵化,直到幼體出來。雄性也可以直接敲打雌性。交配時一般雄性以第一對腹足插入雌性陰道,精子沿腹足中的溝進入雌體受精囊。如雌體是沒有受精囊的種類,則雄性產生精莢,精莢附著在雌體最後兩對胸足的基部,當雌性產卵時,由精莢釋放出精子以受精,並在交配后立刻產卵。少數種類直接產卵於水中,例如對蝦(Penaeus)。但大多數種類產卵於腹部附肢間,並有卵膜物質將卵粘附在一起。多數種類於產卵時受精,受精卵靠附肢的打動造成水流以提供充足的氧。低等的甲殼類可形成卵袋,位於附肢兩側或身體背部,受精卵常變成桔紅色。某些低等的甲殼類行完全卵裂,如鰓足類、橈足類表現出某些螺旋卵裂痕迹。高等的甲殼類均為中黃卵,行表面卵裂,具中空囊胚。發育中有自由游泳的幼蟲,而且不同的種類有不同的幼蟲類型。對蝦最早出現的幼蟲是無節幼蟲(nauplius),身體呈卵圓形,不分節,具三對附肢,第一對為單肢型,后兩對為雙肢型,相當於成體時的第一、二觸角及大顎,具中眼一個。在發育中經連續蛻皮,軀幹體節及相應的附肢逐漸形成,當前8對胸肢形成后稱為溞狀幼蟲(zoea)。但大多數海產蝦類初孵化的幼蟲(第一期幼蟲)為前溞狀幼蟲(protozoea)或溞狀幼蟲,其無節幼蟲期在卵中度過。當所有的附肢獲得后稱為後幼蟲期(postlarva)。后幼蟲期在外形上可能與成體相同,也可能不相同。例如蟹的后幼蟲期腹部仍很長,位於體后並未折回到胸部下面。后幼蟲經蛻皮后完全具備成蟲特徵,只是性器官未成熟。在此基礎上,不同種類的甲殼類可有不同程度的變化。例如對蝦的發育主要經過無節幼蟲、前溞狀幼蟲、糠蝦幼蟲(相當於溞狀幼蟲)及長眼柄幼蟲(后幼蟲)。一些陸生或半陸生種類,繁殖時常返回淡水或海水中以完成其發育。甲殼類的成蟲仍然可以蛻皮,但蛻皮的次數少於幼蟲期。少數種類成蟲后不再蛻皮,例如蟹類。
激素及其調節在無脊椎動物中甲殼綱與昆蟲綱的激素及其調節作用是研究得最清楚的。這裡所說的激素是指內激素(hormone)。外激素(pheromone),也稱信息素,將在昆蟲綱中介紹。激素是由內分泌腺體(endocrine gland)或神經分泌細胞(neurosecretory cells)所分泌的微量物質,直接送入血液或淋巴液,起調節動物生長發育的作用。甲殼類的激素分泌腺體包括眼柄的視神經節內的幾叢神經分泌細胞稱為X器官,它分泌蛻皮抑制激素(moultinhibitinghormone)。X器官的分泌物通過軸突直接運送並貯存在竇腺(sinus gland)內,再由竇腺釋放到血淋巴。另外,在眼柄的腹面及兩側還有神經分泌細胞,稱Y器官,它分泌蛻皮激素(moult hormone),引起甲殼類的蛻皮。完成蛻皮后,X器官分泌的蛻皮抑制激素使之進入蛻皮間期。實驗證明移走Y器官則阻止蛻皮,移走X器官則引起提前蛻皮。引起激素分泌的原因是由於外界刺激作用於中樞的結果,例如光照的長短、溫度的變化等等。甲殼類的生殖及第二性徵也受激素調節。例如,在性成熟之前移走Y器官,則生殖腺的發育嚴重減緩;而在性成熟之後移走,則不受影響。卵巢也與X器官-竇腺系統之間存在著激素調節關係,非生殖期竇腺釋放的激素抑制卵的發育。在生殖季節,中樞神經分泌一種促生殖腺激素,而血液中的生殖腺抑制激素水平下降,卵開始發育。卵巢本身也能產生某種激素,引起第二性徵的出現,例如腹足上攜卵剛毛的出現,卵袋的形成等。雄性精巢及第二性徵的發育也被激素所控制,位於輸精管末端附近的促雄性素腺(androgenic gland)(少數位於精巢內)所產生的激素具有這種作用。移走這一腺體雄性特徵消失,且精巢轉變成卵巢。如將這一腺體植入雌性,則卵巢變精巢,雄性性徵又出現。另外激素也控制著體壁色素細胞的變化,使體色適應外界環境,這在前面已述及。許多甲殼類表現出的某種生理節奏(physiological rhythmic)也是受激素控制的。例如一種綠泳蟹(Carcinus)及招潮蟹,都生活在潮間帶,前者在高潮時活動,後者在低潮時活動,但兩者都通過色素細胞的移動而使體色夜間變淺,白晝時變深暗,表現出色素細胞的節奏移動。甚至將它們飼養在連續光照或連續黑暗的條件下,這一節奏也不改變。但移走眼柄,這一節奏變化消失。相似的節奏如龍蝦(Palinura)的晝夜活動,橈足類的晝夜垂直遷移都是由激素控制的。
自切與再生許多甲殼類,特別是蟹及寄居蟹等經常發生附肢的自切現象(autotomy)。自切的斷面發生在附肢的基節與座節之間的一個雙層膜處,膜上有孔,允許血管、神經通過,並由胸部體壁發出自切肌,直插入到斷裂面。由於外力的牽引或自身因素,自切肌(autotomizer muscle)被刺激而劇烈收縮,由斷裂面處分開,收縮的膜可堵塞血管以防止血液的外流。這種自切現象甚至可由單體節的反射活動而引起。螯蝦僅有螯肢能自切,寄居蟹前三對步足可自切,部分蟹五對步足均可自切。甲殼類也具有一定的再生能力。例如,斷肢后可再生出新肢。之間需經多次蛻皮才能完成,再生部分可以達到丟失部分的大小。演化如同其他節肢動物一樣甲殼亞門的演化過程還不很清楚。這個不清楚的主要原因在於它們的甲殼比較難保存為化石。最早的甲殼亞門化石出現於寒武紀,今天發現的有介形亞綱和軟甲亞綱動物的化石。最早的甲殼亞門動物有可能類似於今天生活在鹽水洞穴中的槳足綱,但它們沒有留下化石。鰓足綱出現於泥盆紀,蔓足亞綱出現於志留紀。尤其介形亞綱動物的殼常常在沉積岩中出現,因此它是重要的指標化石。從它們出現以來它們就是浮遊動物的重要組成部分。在化石中還常出現的有藤壺。
生態意義在海洋和淡水生態系統中甲殼亞門動物,尤其是小的作為浮遊動物的甲殼亞門動物,起了一個非常關鍵的作用。它們食用水中的浮游植物,由此控制這些植物的生長。在浮遊動物中它們所佔的數量比例最大。同時它們也是其他大的水中動物的直接的或間接的食物。人也直接食用一些甲殼亞門動物(比如蝦、螃蟹和龍蝦等)。甲殼亞門也被用來凈化用水,它們有過濾塵埃、細菌和單細胞生物,結合毒質等作用。此外,吸附在船體上的藤壺可以加大船在水中的阻力,帶來經濟損失。
包含分類甲殼亞門分為6個綱(Martin&Davis,2001)。為敘述方便常將小型甲殼動物(前5個綱)統稱切甲類(Entomostracans),將大型的(軟甲綱)稱為軟甲類(Malacostracans)。但這種命名不具分類意義。六綱為:鰓足綱槳足綱頭蝦綱顎足綱介形綱軟甲綱