光周期
光周期
光周期是指晝夜周期中光照期和暗期長短的交替變化。光周期現象是生物對晝夜光暗循環格局的反應。大多數一年生植物的開花決定於每日日照時間的長短。除開花外,塊根、塊莖的形成,葉的脫落和芽的休眠等也受到光周期(指一天中白晝與黑夜的相對長度)的控制。植物對周期性的、特別是晝夜間的光暗變化及光暗時間長短的生理響應特點。尤指某些植物要求經歷一定的光周期才能形成花芽的現象。但其他生理活動也受光周期影響。
人們早就注意到許多植物的開花具有明顯的季節性,同一植物品種在同一地區種植時,儘管在不同時間播種,但開花期都差不多;同一品種在不同緯度地區種植時,開花期表現有規律的變化。
光周期示意圖
,但在冬季轉入溫室栽培后,其株高不足1m就可開花。他們試驗了溫度、光質、營養等各種條件,發現日照長度是影響煙草開花的關鍵因素。在夏季用黑布遮蓋,人為縮短日照長度,煙草就能開花;冬季在溫室內用人工光照延長日照長度,則煙草保持營養狀態而不開花。由此他們得出結論,短日照是這種煙草開花的關鍵條件。後來的大量實驗也證明,許多植物的開花與晝夜的相對長度即光周期有關,即這些植物必須經過一定時間的適宜光周期后才能開花,否則就一直處於營養生長狀態。光周期的發現,使人們認識到光不但為植物光合作用提供能量,而且還作為環境信號調節著植物的發育過程,尤其是對成花誘導起著重要的作用。
人類早已注意到多種植物的開花時間相對穩定,但光周期在決定開花期方面所起的作用直到20世紀才了解清楚。
1912年法國J.圖爾努瓦發現大麻,在每日6小時的短日照條件下會開花,在長日照下則停留於營養生長階段。1913年德國G.A.克萊布斯發現人工加長每日照光時間,可使通常在6月開花的長春花屬(Sem-pervivum)植物能在冬季開花。但明確地提出光周期理論的是美國園藝學家W.W.迦納與H.A.阿拉德。他們在1920年發現,將在美國南部正常開花的煙草(Nicotia-natabacumcv.MarylandMammoth品種)移至美國北部栽培時,夏季只長葉不開花;但如果在秋冬移入溫室則可開花結實。
在北方夏季用遮光辦法縮短日照時數到每天14小時以下,也可使它開花。以後發現大豆(Biloxi品種)、紫蘇、高粱等也有這種現象,並各有其日長上限,日照長度短於此數值時即可開花,稱此日長限度為臨界日長。同時發現菠菜、蘿蔔等植物相反,須在日照長度超過一臨界日長時才能開花。
需光種子需經長日照或連續光處理才能萌發,而嫌光種子則在短光照下與在黑暗中一樣可以萌發。促進或抑制兩類種子的光質要求與成花誘導的要求,都通過光敏素實現。有些植物的營養性貯藏器官的形成,也受光周期的影響。如短日照有利於馬鈴薯形成塊莖,樹木葉片的衰老脫落也與日長有關,一般在秋天短日照下容易脫落。路燈旁的行道樹由於晝夜受光,落葉往往延遲。
人們通過用人工延長或縮短光照的方法,廣泛地探測了各種植物開花對日照長度的反應,發現植物開花對日照長度的反應有以下幾種類型:
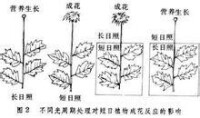
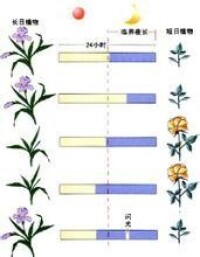
指在24h晝夜周期中,日照長度長於一定時數,才能成花的植物。對這些植物延長光照可促進或提早開花,相反,如延長黑暗則推遲開花或不能成花。屬於長日植物的有:小麥、大麥、黑麥、油菜、菠菜、蘿蔔、白菜、甘藍、芹菜、甜菜、胡蘿蔔、金光菊、山茶、杜鵑、桂花、天仙子等。典型的長日植物天仙子必須滿足一定天數的8.5~11.5h日照才能開花,如果日照長度短於8.5h它就不能開花。
指在24h晝夜周期中,日照長度短於一定時數才能成花的植物。對這些植物適當延長黑暗或縮短光照可促進或提早開花,相反,如延長日照則推遲開花或不能成花。屬於短日植物的有:水稻、玉米、大豆、高粱、蒼耳、紫蘇、大麻、黃麻、草莓、煙草、菊花、秋海棠、臘梅、日本牽牛等。如菊花須滿足少於10h的日照才能開花。
除了以上三種典型的光周期反應類型以外,還有一些其他類型:
中日照植物
光周期
日照條件下才能開花,而在較長或較短日照下均保持營養生長狀態的植物,如甘蔗的成花要求每天有11.5~12.5h日照。
兩極光周期植物
(amphophotoperiodismplant)與中日照植物相反,這類植物在中等日照條件下保持營養生長狀態,而在較長或較短日照下才開花,如狗尾草等。
許多植物成花有明確的極限日照長度,即臨界日長(criticaldaylength)。長日植物的開花,需要長於某一臨界日長;而短日植物則要求短於某一臨界日長,這些植物稱絕對長日植物或絕對短日植物。但是,還有許多植物的開花對日照長度的反應並不十分嚴格,它們在不適宜的光周期條件下,經過相當長的時間,也能或多或少的開花,這些植物稱為相對長日植物或相對短日植物。可以看出,長日植物的臨界日長不一定都長於短日植物;而短日植物的臨界日長也不一定短於長日植物。如一種短日植物大豆的臨界日長為14h,若日照長度不超過此臨界值就能開花。一種長日植物冬小麥的臨界日長為12h,當日照長度超過此臨界值時才開花。將此兩種植物都放在13h的日照長度條件下,它們都開花。因此,重要的不是它們所受光照時數的絕對值,而是在於超過還是短於其臨界日長。同種植物的不同品種對日照的要求可以不同,如煙草中有些品種為短日性的,有些為長日性的,還有些只為日中性的。通常早熟品種為長日或日中性植物,晚熟品種為短日植物。
在溫帶,溫度的季節性變化幅度很大,隨緯度增高而冬季延長。植物如不能在低溫、霜凍到來之前形成種子,就不能繁殖後代。許多植物在長期適應過程中形成了以受氣候條件影響較小的晝夜長短來感應季節的本領。例如一些起源於高緯度地區的植物,常在春季轉暖之後,日照由短變長的條件下開花結實,成花要求長日照。低緯度起源的一些植物,具有短日植物特性,在日長逐漸變短的秋天開花,有利於充分利用低緯度較長的溫暖時期。
在一些不是溫度而是水分對植物生長起決定作用的地區,在冬季為旱季,夏季為濕季的地區起源的植物,屬短日照性的較多,情況相反的地區則多為長日照植物。在同一緯度地區,對日長的不同響應使不同植物在不同季節開花結實,成為決定植物物候期的重要因素。
植物的光周期現象是指植物的花芽分化、開花、結實、分枝習
性、某些地下器官(塊莖、塊根、球莖、鱗莖、塊莖等)的形成受光周期(即每天日照長短)的影響而言。
所謂光周期是指一天中,日出至日落的理論日照時數,而不是實際有陽光的時數。理論日照時數與該地的緯度有關,實際日照時數還受降雨頻率及雲霧多少的影響。在北半球,緯度越高,夏季日照越長,而冬季日照越短。因此,我國北方各地一年中的日照時數在季節間相差較大,在南方各地相差較小。如哈爾濱冬季每天日照只有8—9小時,夏季可達15.6小時,相差6.6—7.6小時。而廣州冬季的日照時數10—11小時,夏季為13.3小時,相差2.3—3.3小時。各地生長季節特別是由營養生長向生殖生長轉移之前,日照時數長短對各類藥用植物的發育是個重要的因素。
植物對光周期的反應通常分為長日照植物、短日照植物、中間型日照植物三類。
長日照植物日照長度必須大於某一臨界日長(一般為12—14小時以上),或者說暗期必須短於一定時數才能形成花芽,否則,植株就停留在營養生長階段。屬於這類的藥用植物有紅花、當歸、莨菪、大蔥、大蒜、芥菜、蘿蔔等。
短日照植物日照長度只有短於其所要求的臨界日長(一般在12—14小時以下),或者說暗期超過一定時數才能開花。如果處於長日照條件下,則只能進行營養生長而不能開花。屬於這類的藥用植物有紫蘇、菊花、蒼耳、大麻、龍膽、扁豆、牽牛花等。
菊花實驗
長度範圍內才能開花結實。而日照長些或短些都不能開花。如野生菜豆只能在每天12一16小時的光照條件下才能開花。又如甘蔗的某些品種,它們只能在日照12小時45分鐘條件下才開花。
應當指出,植物對日照長度的反應是由營養生長向生殖生長轉化的必要條件,是要在其自身發育到一定的生理年齡時才能開始,並非植物的一生都要求這樣的日照長度,而絕大多數藥用植物也絕不是只有一、二次的光周期處理就能引起花芽原基的分化,一般要有十幾次或更多的光周期處理才能引起開花的。
臨界日長是區別長日照或短日照的日照長度的標準,是指每天24小時內光照時間的多少而言,一般為12—14小時。短日照植物要求的日照時數必須短於臨界日長,長日照植物要求的日照時數必須大於臨界日長,否則就不能形成花芽或開花。確切的說,短日照植物並不要求較短的日照,而是要求較長的黑暗,黑暗期的長短對其花芽的形成和開花的影響更為重要。而長日照植物,光照是重要的,黑暗是不重要的,甚至是不必要的。藥用植物的種類很多,對光照長短的反應差異很大,長日照與短日照之間的臨界時數互相交差的情況也是可見的。
植物的光周期反應要在有一定的溫度、植株的生長狀態和營養條件等環境下才能進行正常發育。在影響光周期效應中,溫度是個重要因素。例如,蘿r、、芥菜等長日照植物,將其播種在高溫長日照環境中,它們仍不能開花。這是因為高溫足以抑制長日照對發育的影響。因此從事生產時,必須把光周期與溫度結合起來。同理,還要與植株生長狀態、營養環境等因素結合起來。

光周期
、每天的日照時數等也不盡相同,因此同一藥用植物的生長發育進程也不一致,也就是說,同一藥用植物在各地進入光周期的早晚或通過光周期時間長短也不相同。一般短日照植物在低緯度時進入光周期早,通過的時間也稍短,而長日照植物在高緯度下,進入光周期早,通過時間也快。另外,短日照植物,從北向南引種時,營養生長期縮短,開花結實提前。在作物、蔬菜栽培中,常利用這一特性將北方短日照品種送到南方夏種秋收,爭得一茬收成。
我們的祖先在種菜、種葯及其它作物上,很早就懂得通過改變播期,調整植物生長發育時期的日照條件和溫度條件,達到抑制或促進生長發育的目的。如春播蘿蔔(中藥萊菔子)采籽,秋播蘿I、收肉質根。光周期不僅影響藥用植物花芽的分化與開花,同時也影響藥用植物營養器官的形成。如慈姑、荸薺球莖的形成,都要求較短的日照條件,而洋蔥、大蒜鱗莖的形成要求有較長的光照條件。另外,象豇豆、赤小豆的分枝、結果習性也受光周期的影響……等。
感受光周期的部位
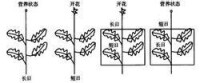
實驗證實:感受光周期反應的部位是植物的葉片:例如用短日照的植物-菊花進行實驗,首先將植株頂端的葉子全部去掉。
實驗表明:葉片感受光周期作用的能力和年齡有關:成熟葉片大於未成熟葉片;不同植物對光周期作用敏感的年齡不同:大多在子葉伸張時期;水稻在七葉期前後過了敏感期之後,年齡越大,光周期誘導的時間越長。
開花刺激物的傳導
接受光周期誘導的部位是葉片,進行光周期反應的部位是莖尖的生長點,葉和起反應的部位之間隔著葉柄和一段莖。那麼,必然有一個開花刺激物傳導的問題。以蒼耳的嫁接實驗來說明。把五株蒼耳植物互相嫁接在一起,且只讓其中一株上的一張葉片處於蒼耳開花適宜的光周期(短日照)下,其他植株都處於不適宜的光周期(長日照)下,它們都可以開花。
這就證明,植株之間確有開花刺激物質通過嫁接的癒合而傳遞。
另外,經過短日照處理的短日植物,例如高冷盤,把其嫁接到長日植物八寶植株上,可引起八寶在短日條件下開花。反之,若將長日處理的長日植物,接到短日植物上,可引起短日植物在長日條件下開花。這說明兩種光周期反應的植株所產生的開花刺激物幾乎具有相同的性質。用蒸汽或麻醉劑處理葉柄或莖,可以阻止開花刺激物的運輸,說明運輸途徑是韌皮部。蘇聯的柴拉軒將這種刺激物叫做成花素(forigen),但這種物質至今還沒有被分離出來。
光周期誘導
光周期誘導:植物只要沒得到足夠日數的適合光周期,以後即便置於不適合的光周期條件下仍可開花,這種現象稱作光周期誘導。

4種類型
物而異:
蒼耳:一個光周期。即15小時照光及9小時的黑暗(15L-9D)。日本短牽牛:一天。大部分短日植物的誘導期需要一天以上,如大豆3天,大麻4天,紅葉紫蘇7-9天,菊12天等。長日植物需要一天的有:白芥、菠菜、油菜、毒素等。一天以上的有:天仙子:2-3天,擬南菜4天,一年生甜菜13-15天等。
不同植物光周期誘導需要的天數與植物年齡、溫度、光照強度、光照長度有關。植物年齡小(達到光周期誘導的能力)、溫度高、光照強,誘導期縮短。
①光周期誘導的光照強度:在自然條件下,光周期誘導所要求的光照強度弱的,遠遠低於光和作用所需的光強度。一般認為在50~100勒克斯之間,有些植物甚至更低,例如水稻在夜間補充光照時,光強只需8~10勒克斯,就能明顯地刺激光周期反應。說明植物光周期反應對光是極敏感的。②暗期光中斷現象和生理效應暗期光中斷現象表明了光周期反應與光強、光質以及參與光反應物質之間的關係。
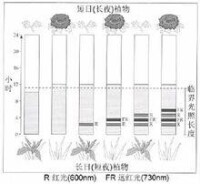
上圖結果表明:暗期對植物的開花更為重要,對於短日植物,它的開花決定於暗期的長度,只要暗期超過臨界夜長(臨界暗期),不管光期多長,它都開花。所以,又稱短日植物為“長夜植物”更為確切。長日植物則相反,它不需要連續黑暗。假如在足以引起短日植物開花的暗期中間,被一個一定強度的閃光所間斷,短日植物就不能開花,長日植物就能開花,這種現象叫做暗期光中斷現象。
用不同波長的光來間斷暗期的試驗表明:無論是抑制短日植物開花,還是誘導長日植物開花,都是紅光最有效。如果在紅光照過後立即再用遠紅光照,那麼暗期閃光間斷的效應就會消失。說明暗期閃光間斷效應有光敏素參與。
光敏素及其在成花中的作用
暗期閃光中斷效應作用的最大作用光譜正好是光敏素的最大吸收光譜,最小作用光譜也正好是光敏素的最小吸收光譜。①光敏素的物理化學性質
光敏素可以從高等植物幾乎所有部分中提取出來,也就是說存在於高等植物的所有組織(根、莖、葉、花、果、實、種子、胚芽鞘)中。在細胞中,光敏素可能集中在細胞的膜的表面上。綠色組織中光敏素含量較低,在黃化組織中較高,濃度級為10-7—10-5M。
高純度的光敏素已從黃化的單子葉植物幼苗(玉米等)中制的。經鑒定是一種藍色蛋白質,它的生色團類似葉綠素和血紅素,有四個吡咯環,但它們不是環狀連接,而是開放
成直鏈,以共價鍵與蛋白質部分相連,即由蛋白質和生色團兩部分所組成。天然的光敏素分子量大約為120kDa。
②光敏素與開花誘導
光敏素有兩種存在形式,Pr和Pfr。在黃化組織中,光敏素大部分以紅光吸收型(Pr)存在,其吸收高峰在660nm。當用紅光照射時,Pr的吸收光譜發生變化,吸收高峰在725nm。這說明通過紅光照射Pr已轉變為另一種形式(Pfr)。
光敏素在開花中如何產生生理效應,當前還不清楚。光敏素本身並不是開花刺激物,但它可以觸發開花刺激物的形成(合成或激活)。當前一般認為,不論是短日植物(SDP)還是長日植物(LDP),其開花都與Pfr與Pr的比例有關。對於SDP,在光期結束時,Pfr/Pr比值高(因為在白天,紅光比例大,有利於Pfr的形成),開花刺激物的合成受到阻止。轉入夜間后,Pfr向Pr逆轉,Pfr/Pr比值變小。當此值到一定水平時,就會觸發引導開花刺激物形成的代謝過程,SDP的成花反應就可以發生。如暗期為紅光所間斷,Pr轉換成Pfr,Pfr/Pr比值升高,開花刺激物的形成即遭受阻止。
對於LDP,形成開花刺激物,要求較高的Pfr/Pr比值,這一比值可在長光期結束時獲得。如果在短日照下,暗期過長,則Pfr轉變為Pr,或Pfr受到破壞,Pfr/Pr值達不到較高水平,開花刺激物就不能形成。暗期的紅光間斷,可使Pfr/Pr值重新提高,使開花刺激物得以合成,長日植物就可以開花。
光周期刺激的感受和傳遞
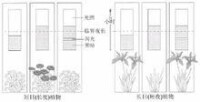
光周期
生長點,然而感受光周期的部位卻是植物的葉片。若將短日植物菊花全株置於長日照條件下,則不開花而保持營養生長;置於短日照條件下,可開花;葉片處於短日照條件下而莖頂端給予長日照,可開花;葉片處於長日照條件下而莖頂端給予短日照,卻不能開花(圖9-10)。這個實驗充分說明:植物感受光周期的部位是葉片。對於光周期敏感的植物,只有葉片處於適宜的光周期條件下,才能誘導開花,而與頂端的芽所處的光周期條件無關。雖然也有少數植物的其它部位對光周期有一定的敏感性,如組織培養的菊苣根可對光周期起反應,但感受光周期最有效的部位是葉片。葉片對光周期的敏感性與葉片的發育程度有關。幼小的和衰老的葉片敏感性差,葉片長至最大時敏感性最高,這時甚至葉片的很小一部分處在適宜的光周期下就可誘導開花。例如,蒼耳或毒麥的葉片完全展開達最大面積時,僅對2cm2的葉片進行短日照處理,即可導致花的發端。
對昆蟲滯育產生影響的光周期(Photoperiod)是指一晝夜中的光照時數與黑暗時數的節律,一般以光照時數表示,是影響昆蟲滯育的主要因素。引起昆蟲種群中50%的個體滯育的光照時數,稱為臨界光周期(critical photoperiod)。不同種或同種不同地理種群的昆蟲,其臨界光周期不同,如三化螟南京種群為13小時30分,廣州種群為12小時。感受光照刺激的蟲態,稱為臨界光照蟲態。臨界光照蟲態常常是滯育蟲態的前一蟲態,如家蠶以卵滯育,其臨界光照蟲態為上一代成蟲;亞洲玉米螟以老熟幼蟲滯育,其臨界光照蟲態為3~4齡幼蟲;棉鈴蟲以蛹滯育,其臨界光照蟲態為1~5齡幼蟲。處於臨界光照蟲態的昆蟲對光的反應極為敏感,一般只需l~2Lx(勒克斯)的照度就能發生作用,如鑽入蘋果內的梨小食心蟲,當果面的光照度只有1~3Lx時,即能對光作出反應。根據昆蟲滯育對光周期的反應,可將昆蟲分為以下4種滯育類型。
1、短日照滯育型 即長日照發育型。其特點是昆蟲滯育的個體數隨日照時數的減少而增多。通常光周期長於12~16小時,仍可繼續發育而不滯育。一般冬季滯育的昆蟲,如亞洲玉米螟等屬於此類型。
2、長日照滯育型 即短日照發育型。其特點是昆蟲滯育的個體數隨日照時數的增加而增多。通常光周期短於12小時,仍可繼續發育而不滯育。一些夏季滯育的昆蟲,如小麥吸漿蟲等屬於此類型。
3、中間型光周期過短或過長均可引起滯育,只有在相當窄的光周期範圍內才不滯 育。如桃小食心蟲在25℃時,光照短於13小時,老熟幼蟲全部滯育;光照長於17小時,半數以上滯育;而光照為15小時時,則大部分不滯育。
4、無光周期反應型 光周期變化對滯育沒有影響。如蘋果舞毒蛾、丁香天蛾等。
光周期
短日(照)植物
要求經歷一段白晝短於一定長度、黑夜長於一定長度(稱為臨界暗期)的時期才能開花的植物。延長暗期有利於成花。這一類植物多半起源於低緯度熱帶地區,也包括一些在溫帶地區早春或晚秋開花的庭園花卉和野花。如美洲煙草、大豆、水稻、玉米、粟、菊花、蒼耳等。
長日(照)植物
要求經歷一段白晝長於一定的臨界值(臨界日長)、黑夜短於一定長度的時期才能成花的植物。延長光照時間、縮短黑暗時間,可以提早成花。這類植物包括許多溫帶起源的作物、晚春與初夏開花的花卉或野花。如大麥、小麥、黑麥、蘿蔔、菠菜、甘藍、大白菜、天仙子、甜菜等。
中(間)日(照)性植物
只有在某一範圍的日長下才能成花的植物。日照過短或過長,均不利於開花。這類植物種類很少。
日照中性植物
也稱日長鈍感植物。成花對晝夜長短無一定要求,也無明顯響應。這類植物種類最多。如棉花、番茄、茄子、四季豆和一些四季開花的花卉(月季)等。
此外,還有一些植物要求先經歷一段短日照時期,再經歷一段長日照時期,才能開花,如風鈴草等。
另一些植物要求先經歷長日照時期,再經歷短日照時期,才能開花,如落地生根等。
光周期應用
光敏素
掌握和控制其開花、結實等生長發育時期在一年中的分配,具有重要意義。例如:
①引種時要詳細了解當地日長的季節變化和待引品種的光周期要求。如果日長條件不符合其要求,則不能開花或開花延遲,種子不能成熟;過早地滿足成花要求,又會不必要地縮短生長季,對干物質的累積不利。
②人工縮短或延長光照時間可以促進或延遲開花,甚至可以使許多原來開花季節較固定的花卉在任何季節開花。在雜交育種中可以用人工長日或短日處理來使父母本花期相遇,提高雜交種子產量。
③在育種工作中,當作物種子在收穫后立即播種,而光周期不利於開花結實時,人工調整日長,可使之及時成花、結實,從而加速世代繁育,縮短育種年限。中國育種學家創造了南繁北育的辦法,利用異地溫度和光周期的條件,使某些作物可一年繁育2~3代。例如:中國水稻育種家常利用海南島冬季日照短、較溫暖來進行冬繁,使良種繁育時期縮短。
移至北方栽培,利用長日條件推遲或防止成花,可提高煙葉或麻纖維的產量或品質。
利用對植物光周期特性的知識控制生長發育的辦法,大體上可分為兩類:
①人工改變日長。因為成本較高,多用於經濟價值較高的花卉的栽培或作物品種的繁育。其中光間斷打斷長夜所需光量較小,比人工照光延長光照時間經濟得多。
②選擇品種特性,以適應特定地區自然光照特點。這可用於多種作物。為了使所得品種能推廣於較大的緯度範圍,近代育種工作者有選育對光周期較不敏感的遺傳特性的趨勢。
光周期在農業生產上的應用

光周期
的影響較為突出,南北距離較遠時,則不易成功。長日照植物南北引種,光溫影響是補償的,一般較易成功。但在熱量條件較差地區,從高緯度引種短日照植物,往往有利於避霜早熟。但對收穫營養體為主的作物,則要防止過早向生殖生長轉化。
2、育種工作中利用光周期調節花期實現品種間雜交。利用光周期效應使作物提早發育,還可以實現加代繁殖縮短育種年限。
3、調節播栽期。選擇適宜的播種期和移栽期,使自然日照、溫度與作物的要求相一致,是提高產量的主要條件。
4、花卉栽培。利用光周期現象進行人工調節花期,能使短日照植物的菊類在任何季節開花。
光周期
在可見光中,光照誘導成花或光間斷破壞誘導的效應,以660納米的紅光為最有效。這種作用可以被730納米的遠紅光解除,又可以被660納米的紅光恢復。這種相互逆轉可以反覆多次。這表明接受光周期信號的色素是植物。660納米的紅光將光敏素轉變為生理上活躍的遠紅光型,而730納米的遠紅光則把遠紅光型轉變為生理上不活躍的紅光型(Pr)。在黑暗中的衰變速率和向Pr的轉化速率,可能對光周期響應的計時起一些作用。但的暗衰敗速率隨溫度升降變化很明顯,而有些植物的光周期長短卻突出地不受溫度變化的影響,所以植物體內必定還另有確定節奏的生物鐘。它的機理,至今仍不清楚。有一些植物的光周期響應在高溫下與低溫下不同。例如短日植物中的牽牛、高冷盤、蒼耳等,在低溫條件下,即使在長日照下也可成花。長日植物中的二色金光菊(Rudbeckiabicolor)、風輪菜屬植物(Calaminthaofficinalis)在高溫處理下,Blitumcapitatum與B.viratum在低溫下均可在短日照下開花。
光周期誘導成花過程在解剖上最早看到的是苗端開始分化花芽,但是否分化花芽取決於葉片是否處於適當的光周期條件下。可見感受光周期的部位不是苗端,而是葉片。有些對光周期敏感的植物,只要一片葉或甚至一片葉的一部分(例如蒼耳只要長成葉的2平方厘米)接受合適的光周期,即使其餘部分都處在不利於成花的光周期下,也能分化花芽。顯然,接受光周期的葉片,必須以某種方式把光周期誘導的信號傳遞給苗端的分生組織。
葉片所接受的光周期刺激,不但可以傳遞給本枝條,而且在本枝條去頂芽后,還可以傳遞給同株的另一個去掉了葉片的枝條的頂芽,使它分化出花芽。當把經過誘導的植株的葉片嫁接到另一株未經誘導的植株上去以後,也可以誘導它成花。從后一植株取下的葉片,又可以嫁接的方式誘導另一株未經誘導的植株成花。這種嫁接在蒼耳上可以輾轉5~6次,仍然生效。
根據這類實驗結果,蘇聯M.X.柴拉希揚於1937年提出,誘導葉中形成一種物質,傳導到頂芽分生組織后引起成花,他稱這種物質為成花素。但經過幾十年的努力,還未能分離和鑒定出這種物質。
另一些植物,如草莓、藜(短日植物)和天仙子(長日植物),只要將全部葉子去掉(新葉隨生隨除),在任何日長下都能開花。表明在這些植物的葉子里,不是在適宜的日照下形成促進花芽分化的物質,而是在不適宜的日照下形成某種抑制花芽分化的物質,稱為成花抑制物,所以去掉葉片反而容易開花。
無論長日植物或短日植物,一般只要求經歷一定天數的長日或短日,就能成花,並不要求長期處於那種日照長度之下。有些短日植物,如蒼耳,甚至一個短日長夜周期,即可成花。但從這一合宜的光周期處理到花芽分化,還需一段時期。這種適合的光暗周期使植物從不分化花芽的營養生長階段轉入分化花芽的狀態的作用,稱為光周期誘導。
長日植物所需的日照長度因植物種類而異。如一年生甜菜為13~14小時,冬小麥為12小時,天仙子為10小時,並不一定要比12小時長。短日植物所需的臨界日長多數短於12小時,但也有長於12小時的,如某些大豆品種為13.5小時,蒼耳為15.5小時。所以在晝長13小時夜長11小時的光周期下,長日植物冬小麥、天仙子和短日植物晚大豆(Biloxi)、蒼耳都可以開花。可見,長日植物與短日植物的區別,不在於臨界日長是否大於12小時,而在於要求日長大於還是小於某臨界日長。
無論對於短日植物或長日植物,以光將長夜打斷,都產生類似於短夜的效應(短日植物不能成花,長日植物能成花);而以暗間斷將長日打斷,卻不影響長日的效應(短日植物仍不能成花,長日植物仍能成花)。所以關鍵是夜長而非晝長。但夜長也不是決定能否誘導成花的唯一因素。如只需一天短日長夜的藜,在連續光照條件下,給以72小時黑暗處理,並在這個暗期內的不同時間做光間斷處理,如果在暗期開始3小時后照射2分鐘紅光,則雖然以後的暗期還有69小時,遠遠大於臨界暗期,植物也不能成花。而如果在暗期開始后18小時做光間斷處理,則植株不但能開花,而且成花數比一個通常的短日長夜處理還多。光間斷效應的大小與正負,與它在光周期中的時刻有關。按E.比寧的理論,許多植物(以及動物)體內有一種內在節奏,稱為生物鐘或生理鍾。它的周期接近但不等於24小時,一般在21與28小時之間。這個周期稱為近似晝夜周期。在周期的不同相位上,植物對光或暗處理的敏感性不同。