水解纖維素
水解纖維素
纖維素水解的產物稱為水解纖維素,它不是一種有固定組成的化合物,而是一種依水解程度不同而不同的混合物,纖維素完全水解即為葡萄糖。水解纖維素主要是由於纖維素在酸、酶等因素的作用下產生的,其化學結構與原纖維素相同,主要區別在於它的聚合度小於原纖維素,因此,水解纖維素的機械強度大大低於原纖維素。
1906年,Seilliere首次在蝸牛的消化液中發現了能分解天然纖維素的纖維素酶。1945年又在微生物中發現了此酶。到了20世紀六七十年代,科學家們針對世界人口猛增的形勢,開始研究用纖維素酶使纖維索轉化為食物,生產單細胞蛋白。第二次世界大戰后,前蘇聯每年用於飼料的單細胞蛋白高達200萬噸。進人20世紀90年代,世界範圍的能源枯竭和環境污染日益嚴重,纖維素酶的研究重點又轉變為開闢新能源及防止廢纖維污染。有人將纖維索酶的研究分為三個階段(閻伯旭等,1999):第一階段是20世紀80年代前,主要利用生物化學的方法對其進行分離純化;第二階段是1980- 1988年期間,主要利用基因工程的方法對酶的基因進行克隆和一級結構的測定;第三階段是1988年至今,主要利用結構生物學及蛋白質工程的方法對纖維索酶分子結構及功能進行研究。
纖維素酶類作為飼用添加劑,國外從20世紀70年代起開始對其進行較為系統的研究。由於纖維素酶的活性不高,用最過大,來源有限,致使生產成本過高而應用受到限制。20世紀80年代後期,由於酶的生產技術和菌種篩選、分子生物學的發展及生物技術取得了突破性進展,酶的活力單位提高,單位酶活力的生產成本不斷下降,纖維素酶的應用研究才得以迅速展開,從而使其在飼料中的應用出現了新的前景。
纖維素酶是能將纖維素水解成葡萄糖的一組酶的總稱,來源廣泛,主要包括微生物來源、植物來源和動物來源纖維素酶。
1、微生物來源的纖維素酶
能分泌纖維素酶的微生物主要有黴菌、擔子菌等真菌,也包括細菌、放線菌和一些原生動物。目前,人們研究的纖維素酶主要來自於細菌和絲狀真菌(Wood,1992;Bhat和Bhat,1997)。細菌主要有纖維黏菌、生孢纖維黏菌和纖維桿菌等,但由於細菌分泌的纖維素酶量少(低於0.1 g/L),同時產生的酶屬胞內酶或者吸附於細胞壁上,故很少用細菌作纖維素酶的生產菌種。絲狀真菌則能較大量地產生纖維素酶,且能分泌到細胞外,屬胞外酶,這有利於酶的提取。
利用微生物生產纖維素酶的研究開展較早。早期許多研究集中在利用綠色木霉(、康氏木霉、青黴等嗜溫好氧真菌產纖維素酶。對纖維素作用較強的菌株多是木霉屬、青黴屬、麴黴屬和枝頂孢霉的菌株,特別是綠色木霉及其近緣的菌株。目前飼用纖維素酶主要來源於綠色木霉、李氏木霉、根霉、青黴、嗜纖細菌、側孢菌等,其中綠色木霉應用最為廣泛。
2、植物來源的纖維素酶
植物可以產生纖維素酶的觀點早已被人們所認識和接受。在植物中,纖維素酶在植物發育的不同階段發揮著水解細胞壁的作用,如果實成熟、蒂柄脫落等過程。柴國花等(2006)採用RT-PCR檢測培養4周的大豆幼苗的5個不同組織:嫩葉、老葉、莖、離層和根,測得脫落纖維素酶基因的表達量互不相同,離層中表達量最高,莖中表達量最低。同時選取表達量最高的離層作為逆境處理材料,分別用高溫、乾旱、鹽處理不同時問后,檢測脫落纖維素酶基因的時間表達模式,結果表明:3種逆境條件下,脫落纖維素酶基因的時間表達模式各不相同,但總的來說,高溫能抑制脫落纖維素酶基因的表達,乾旱和鹽都能促進脫落纖維素酶基因的表達。目前關於穀物性飼料中是否存在纖維素酶及其對畜禽消化影響的研究並不多見.對植物源纖維素酶的酶活及其調控研究將有助於研究植物源和微生物源纖維素酶的差異並進一步開發高效纖維素酶菌種。
3、動物米源的纖維素酶
動物來源纖維素酶包括在動物消化道內寄生微生物分泌的纖維素酶和動物自身分泌的纖維素酶。反芻動物依靠瘤胃微生物可消化纖維素,因此可以利用瘤胃液獲得纖維素酶的粗酶製劑。張曉華等(1995)報道了一個厭氧中溫分解纖維素的瘤胃梭菌新種。Nakashima等(2000)從白蟻體內分離到一種相對分子質量為48000的內切β-1,4-葡聚糖酶。王驥(2003)從福壽螺體內分離得到一種同時具有外切β-1,4-葡聚糖酶、內切β-1,4葡聚糖酶和內切北塔-1,4一木聚糖酶3種酶活性,相對分子質量為41500的多功能纖維素酶,同時在福壽螺的卵母細胞中獲得了編碼該酶的基因。上述研究證實動物自身可分泌內源性纖維素酶,這可能是動物在進化過程中對自然環境的適應性選擇,對高等動物豬、雞等而言自身分泌的纖維素酶有限。
4、液體發酵產纖維素酶
液體發酵的突出優點是便於控制污染,尤其是目前常用的液體深層發酵技術,另外就是產生的纖維素酶純度高、便於濃縮成高濃度的產品。液體發酵節省勞動力、適合於大規模工業化生產,但大規模生產時,發酵罐的攪拌槳不停地攪拌耗能是相當巨大的。液體發酵的培養周期長,至少長達7~8 d,更多的長達11 d以上(丹尼爾等,1984)。
5、固體發酵產纖維素酶
固體發酵的基本流程是:菌種平板一搖瓶一種曲一固體發酵。固體發酵產纖維素酶的特點是:由於發酵條件更接近自然環境狀態下的微生物生長習性,使得其產生的酶系更全面,有利於降解天然纖維素;在設備、耗能、投資、生產成本方面國內外許多丁-廠的建立已說明比液體發酵優越得多;固體發酵不需攪拌,培養周期短,只培養3 d,且在不含遊離水的條件下培養,水分為75%。但是,固體發酵比液體發酵較難控制污染,不過隨著近幾年技術發展,固體發酵污染相對較難控制的問題,已得到很大的改進。
我們在2004年的研究中分別選擇了1種固體發酵纖維素酶和2種液體發酵纖維素酶,均來源於真菌。比較而言,固體發酵獲得CMCase活力較高,但總的FPase反而液體發酵較高;此外,液體發酵來源纖維素酶具有比固體發酵更好的穩定性,表現在抗逆性和熱穩定性等方面(黃燕華,2004)。
關於纖維素酶降解纖維素作用機制的假說很多,在對纖維素酶各組分間作用順序和部位及協同作用的分子學機理等方面的假說存有較大異議,但對於各組分間存有很強的協同作用及部分單一組分酶的水解效果具有一定的共識。
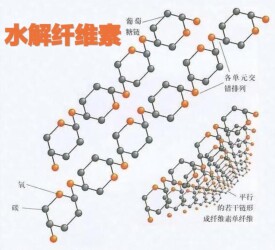
1950年,Reese等提出了由於天然纖維素的特異性而必須以不同的酶協同作用才能分解的C1-Cx假說。這個假說認為:當纖維素酶作用時,首先C1酶(內切葡聚糖酶)首先作用於纖維素結晶區,使其轉變成可被Cx酶(外切葡聚糖酶)作用的非結晶區,Cx酶隨機水解非結晶區纖維素,然後β-1,4一葡萄糖苷酶將纖維二糖水解成葡萄糖。Wood(1985)在研究木霉、青黴的纖維素酶水解纖維素時,發現培養液中的兩種外切酶在液化微晶纖維素和棉纖維時具有協同作用。Kanda等(1976)還發現了只是對可溶性纖維素進攻方式不同的兩種內切葡萄糖苷酶在結晶纖維素的水解過程中也具有協同作用。協同作用一般認為是內切纖維素酶首先進攻纖維素的結晶區,形成外切纖維素酶需要的新的遊離末端。然後外切纖維素酶從多糖鏈的非還原性末端切下纖維二糖單位,β-葡萄糖苷酶再水解纖維二糖單位,形成葡萄糖。一般來說,協同作用與酶解底物的結晶度成正比,當酶組分的混合比例與黴菌發酵濾液中各組分比相近時,協同作用最大,不同菌源的內切與外切酶之間也具有協同作用(Henrissat等,1995)。目前最易接受的酶水解機理如圖所示
纖維素水解圖
近年來,應用凝膠過濾、離子交換層析、凝膠電泳以及等電聚焦等生化分離分析手段,發現纖維素酶非常複雜。研究發現,內切葡聚糖酶、外切葡聚糖酶及β-葡萄糖苷酶均存在2~4個同功酶,對於這些同功酶的功能,目前尚未有明確的闡述。
水生細菌有的具有分解纖維素的能力,通常認為,能在以纖維素作為唯一碳源的培養基上生長的細菌必然含有纖維素酶,即能分解纖維素,如纖維分解細菌等具有分解纖維素的能力。故而可利用此類細菌的這種特性來鑒定它們。鑒定時以不含碳源的培養基為宜,下舉兩種。
1、無機鹽培養基
NHNO 1.0克
KHPO·3HO 0.5克
KHPO 0.5克
MgSO·7HO 0.5克
NaCl 1.0克
CaCl 0.1克
FeCl 0.02克
酵母膏 0.05克
水 1000毫升
將以上成分溶於水,調pH為7.0-7.2。分裝於試管,在121℃下的高壓蒸汽鍋內滅菌20分鐘。
2、腖水培養基
蛋白腖 5克
NaCI 5克
自來水 1000毫升
將上面兩物質放於水中,加熱溶解。調pH為7.0-7.2,分裝於試管,在121℃的高壓蒸汽鍋內滅菌20分鐘。