wnt信號通路
複雜的蛋白質作用網路
Wnt信號通路是一個複雜的蛋白質作用網路,其功能最常見於胚胎髮育和癌症,但也參與成年動物的正常生理過程。 Wnt信號傳導途徑是由配體蛋白質Wnt和膜蛋白受體結合激發的一組多下游通道的信號轉導途徑。經此途徑,通過細胞表面受體胞內段的活化過程將細胞外的信號傳遞到細胞內。Wnt是遺傳學領域的首字母縮寫,代表“Wingless/Integrated”。
Wnt信號途徑在細胞間呈現兩種信號傳導方式:胞間交流(旁分泌)或者自體細胞的交流(自分泌)。Wnt信號通路在動物間存在遺傳學上的高度保守性,不同的動物物種間極為相似。
1982年,Wnt基因首次在小鼠乳腺癌中發現,由於此基因激活依賴小鼠乳腺癌相關病毒基因的插入,因此,當時被命名為Int1基因。之後的研究表明,Int1基因在小鼠正常胚胎髮育中發揮重要作用,與果蠅的無翅(Wingless)基因功能相似,均可控制胚胎的軸向發育。 此後大量研究提示Int1基因在神經系統胚胎髮育中的重要性。因兩者基因與蛋白功能的相似性,研究者將Wingless與Int1合併,賦名為Wnt基因。人Wnt基因定位於12q13。在胚胎髮育中,Wnt基因調控的重要信號傳導系統統稱為Wnt通路。
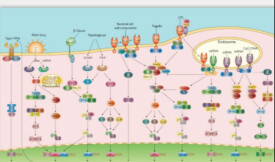
Wnt蛋白(Wnt配體)、Wnt受體(Frizzled家族蛋白及低密度脂蛋白受體相關蛋白(LDLreceptorrelatedprotein,LRP))、Dishevelled(Dsh/Dvl)蛋白、β-連環蛋白(β-catenin)、糖原合成酶激酶3β(GSK-3β)、Axin/Conductin、APC(adenomatouspolyposiscoli)蛋白等。
3、Wnt/Ca通路,激活磷脂酶C(PLC)和蛋白激酶C(PKC);
經典的Wnt途徑(Wnt/β-連環蛋白途徑)導致基因轉錄的調節,並且被認為部分地由SPATS1基因負調節。
Wnt/β-連環蛋白途徑是Wnt途徑中的一種,該途徑會導致β-連環蛋白在細胞質中積累並最終會作為屬於TCF的轉錄因子的轉錄共激活因子/LEF家族易位至細胞核。
沒有Wnt,β-連環蛋白不會在細胞質中積累,因為有一種破壞複合物通常會降解它。該破壞複合物包括以下蛋白質:Axin,腺瘤病大腸桿菌(APC),蛋白磷酸酶2A(PP2A),糖原合成酶激酶3(GSK3)和酪蛋白激酶1α(CK1α)。它通過將β-連環蛋白靶向泛素化而降解β-連環蛋白,隨後將其送至蛋白酶體進行消化。
當Wnt蛋白與Frizzled(Fz)家族受體的N末端細胞富含半胱氨酸的結構域結合時,Wnt信號開始。這些受體跨越質膜七次,構成了一個獨特的G蛋白偶聯受體家族(GPCR)。然而,為了促進Wnt信號傳導,可能需要共同受體以及Wnt蛋白和Fz受體之間的相互作用。例子包括脂蛋白受體相關蛋白(LRP)-5/6,受體酪氨酸激酶(RTK)和ROR2。但是,只要Wnt綁定Fz和LRP5/6,破壞複合物的磷酸化就會被阻斷從而使得其功能中斷。破壞複合物中由其他蛋白引起的磷酸化隨後將Axin與LRP5/6的細胞質尾部結合。Axin去磷酸化了,它的穩定性和濃度水平降低。然後Dsh通過磷酸化被激活並且它的DIX和PDZ結構域抑制GSK3破壞複合物的活性。這使得β-連環蛋白能夠積聚並定位於細胞核,隨後通過基因轉導以及TCF/LEF(T細胞因子/淋巴增強因子)轉錄因子誘導Wnt最終作用的目標基因轉錄,誘導後續的細胞反應的發生。β-連環蛋白募集其他轉錄共激活因子,如BCL9,Pygopus和Parafibromin/Hyrax。多虧有了新的高通量蛋白質組學研究,β-連環蛋白組裝的轉錄複合物的複雜性開始出現。β-連環蛋白相互作用蛋白的混雜性使我們的理解變得複雜:實際上已報道BCL9和Pygopus具有幾種不依賴於β-連環蛋白的功能(因此,可能是獨立於Wnt信號途徑的)。
多細胞生物體軸分化過程中起重要作用
β-連環蛋白調節的典型Wnt信號參與前後軸的形成;
β-連環蛋白敲除的胚胎,可發生細胞的錯誤定位,從而不能形成中胚層;
抑制Wnt信號是脊椎動物體廓形成後期階段的關鍵因素;
Wnt拮抗分子能誘導頭的形成。
Wnt信號與器官發生
Wnt信號參與大腦的形成。Wnt3a敲除的小鼠胚胎,大腦海馬回發育受損;Lef純合子突變可導致小鼠胚胎缺少全部海馬回;Wnt/LEF/TCF基因協同作用,共同參與大腦海馬回的發育。
Wnt信號參與生長錐的重建和多突觸球狀環(苔狀神經纖維與顆粒細胞相接觸時)的形成。參與軸突形成的起始過程:Wnt7a能誘導苔狀神經纖維中軸突和生長錐的重建和觸素Ⅰ的彙集。
Wnt信號參與脊椎動物的肢體起始和頂端外胚層脊的形成。三種Wnt信號分子(Wnt2b、Wnt3a、Wnt8c)是信號轉導的關鍵誘導者;FGF與Wnt信號的信息交流也與內耳的形成有關。
Wnt信號與腫瘤發生
調控EMT的信號通路
參與調控EMT過程的信號通路網路簡介:Wnt信號通路能通過抑製糖原合成酶激酶3β(glycogensynthasekinase-3β,GSK3β)介導的磷酸化作用以及抑制胞質中的β-連環蛋白(β-catenin)降解等作用來誘發EMT轉換。胞內丰度大量增加的β連環蛋白會轉移進入核內,作為轉錄因子亞單位誘導大量基因的表達,這些靶基因的表達產物中有很多都是能夠誘導EMT轉換過程的轉錄因子。
通過基因沉默事件導致細胞依賴於WNT信號通路:Wnt信號通路廣泛存在於無脊椎動物和脊椎動物中,是一類在物種進化過程中高度保守的信號通路。Wnt信號在動物胚胎的早期發育、器官形成、組織再生和其它生理過程中,具有至關重要的作用。如果這條信號通路中的關鍵蛋白髮生突變,導致信號異常活化,就可能誘導癌症的發生。a在正常的結腸上皮細胞中,分泌性的frizzled相關蛋白(SFRPs)的功能是與WNT競爭性地同Wnt受體Frizzled結合,從而拮抗Wnt信號。當Wnt信號失活,腺瘤息肉病基因(APC)複合物磷酸化β-catenin,導致β-catennin降解。這便阻止了β-catenin的核內沉積,則不能激活轉錄因子(TCF),最終導致細胞進入分化並保持結腸上皮細胞處於動態平衡狀態。b通過表觀遺傳調控的基因沉默使得SFRP表達缺失,即“表觀遺傳調控門控基因”缺失,Wnt信號通路激活,促進細胞增殖以及存活而不進入分化。c持續性的激活Wnt信號使得信號通路中的其他分子有可能發生突變,例如永久性失活APC複合物(圖中為粗體*所示),即“遺傳調控門控基因”缺失,進一步激活Wnt信號通路,從而促進腫瘤的發展。
基本信息
- 中文名
- wnt信號通路
- 外文名
- Wnt signal path
- 發現
- 1982年小鼠乳腺癌發現了Wnt基因
- 組成
- Wnt蛋白等
- 分類
- 平面極細胞通路等