蛋白酶體是在真核生物和古菌中普遍存在的,在一些原核生物中也存在的一種巨型蛋白質複合物。在真核生物中,蛋白酶體位於細胞核和細胞質中。蛋白酶體的主要作用是降解細胞不需要的或受到損傷的蛋白質,這一作用是通過打斷肽鍵的化學反應來實現。能夠發揮這一作用的酶被稱為蛋白酶。蛋白酶體是細胞用來調控特定蛋白質和除去錯誤摺疊蛋白質的主要機制。經過蛋白酶體的降解,蛋白質被切割為約7-8個氨基酸長的肽段;這些肽段可以被進一步降解為單個氨基酸分子,然後被用於合成新的蛋白質。需要被降解的蛋白質會先被一個稱為泛素的小型蛋白質所標記(即連接上)。這一標記反應是被泛素連接酶所催化。一旦一個蛋白質被標記上一個泛素分子,就會引發其它連接酶加上更多的泛素分子;這就形成了可以與蛋白酶體結合的“多泛素鏈”,從而將蛋白酶體帶到這一標記的蛋白質上,開始其降解過程。從結構上看,蛋白酶體是一個桶狀的複合物,包括一個由四個堆積在一起的環所組成的“核心”(右圖中藍色部分),核心中空,形成一個空腔。其中,每一個環由七個蛋白質分子組成。中間的兩個環各由七個β亞基組成,並含有六個蛋白酶的活性位點。這些位點位於環的內表面,所以蛋白質必須進入到蛋白酶體的“空腔”中才能夠被降解。外部的兩個環各含有七個α亞基,可以發揮“門”的作用,是蛋白質進入“空腔”中的必由之路。這些α亞基,或者說“門”,是由結合在它們上的“帽”狀結構(即調節顆粒,右圖中紅色部分)進行控制;調節顆粒可以識別連接在蛋白質上的多泛素鏈標籤,並啟動降解過程。包括泛素化和蛋白酶體降解的整個系統被稱為“泛素-蛋白酶體系統”。蛋白酶體降解途徑對於許多細胞進程,包括細胞周期、基因表達的調控、氧化應激反應等,都是必不可少的。2004年諾貝爾化學獎的獲獎主題就是蛋白質酶解在細胞中的重要性和泛素在酶解途徑的作用,而三位獲獎者為阿龍·切哈諾沃、阿夫拉姆·赫什科和歐文·羅斯。
蛋白酶體20S核心顆粒的簡化結構圖。構成外部兩個環的α亞基用綠色來表示,構成中間兩個環的β亞基用藍色來表示。
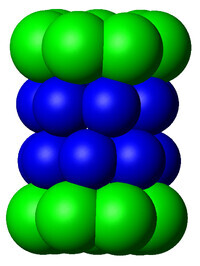
蛋白酶體20S核心顆粒的簡化結構圖。構成外部兩個環的α亞基用綠色來表示,構成中間兩個環的β亞基用藍色來表示。
從上往下看核心顆粒的簡化結構。可以看出環結構存在七次軸對稱。
蛋白酶體的組分通常根據它們的斯維德伯格沉降係數(以“S”來標記)來命名。最普遍的蛋白酶體的形式是26S蛋白酶體,其分子量約為2000kDa,包含有一個20S核心顆粒和兩個19S調節顆粒。核心顆粒為中空結構,將剪切蛋白質的活性位點圍在“洞”中;將核心顆粒的兩端敞開,目的蛋白質就可以進入“洞”中。核心顆粒的每一端都連接著一個19S調節顆粒,每個調節顆粒都含有多個ATP酶活性位點和泛素結合位點;調節顆粒可以識別多泛素化的蛋白質,並將它們傳送到核心顆粒中。除了19S調節顆粒外,還存在另一種調節顆粒,即11S顆粒;11S調節顆粒可以以類似於19S顆粒的方式與核心顆粒結合;11S顆粒可能在降解外源肽(如病毒感染后產生的肽段)上發揮作用。此外,PA200(酵母中為Blm10)蛋白也可以單獨作為激活蛋白來調控20S顆粒的開啟。
不同的生物體中,20S核心顆粒中
亞基的數量和差異性都有所不同;就亞基數量而言,
多細胞生物比單細胞生物要多,真核生物比原核生物多。所有的20S顆粒都由四個堆積的七元環所組成,這些環結構則是由兩種不同的亞基構成:α亞基為結構性蛋白,而β亞基則發揮主要的催化作用。外部的兩個環,每個環都含有七個α亞基,一方面作為調節顆粒的結合部,另一方面發揮“門”的作用,阻止蛋白質不受調控地進入核心顆粒的內部。內部的兩個環,每個環都含有七個β亞基,且包含蛋白酶活性位點,用於蛋白質水解反應。蛋白酶體的大小在不同物種之間相當保守,其長和寬分別為約150 Å和115 Å。其內部孔道寬為近53 Å,而入口處則只有13 Å的寬度,這就提示蛋白質要進入其中,需要先被至少部分去摺疊。
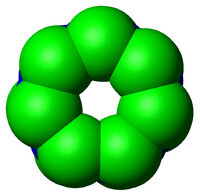
從上往下看核心顆粒的簡化結構。可以看出環結構存在七次軸對稱。
在古菌(如Thermoplasma acidophilum(英語:Thermoplasma))中,所有的α亞基和所有的β亞基是等同的;而真核生物的蛋白酶體(如
酵母)中,每個亞基都不相同,即α和β亞基都含有七種不同的亞基。在哺乳動物中,和亞基具有催化作用;雖然它們有著共同的催化機制,但它們具有不同的底物特異性,分別為類胰凝乳蛋白酶型、類胰蛋白酶型和肽谷氨醯基肽水解(英語:peptidyl-glutamyl peptide-hydrolyzing)。在暴露於前炎症信號(如
細胞因子,特別是γ干擾素)時,細胞應激反應會促使造血細胞表達另一些形式的β亞基,即。由這些替代亞基所組裝成的蛋白酶體又被稱為“免疫蛋白酶體”(immunoproteasome),相對於正常形式的蛋白酶體,其底物特異性發生了變化。
真核生物中的19S顆粒是由19個蛋白質組成的,並可以被分成兩個部分:一個由10個蛋白質組成的可以與20S核心顆粒上的α環直接結合的基底,和一個由9個蛋白質組成的結合多泛素鏈的蓋子。其中,10個基底蛋白質中的6個具有
ATP酶活性。19S和20S顆粒的結合需要ATP先結合到19S顆粒上的ATP結合位點。 ATP的水解對於蛋白酶體降解一個連接泛素的緊密摺疊的蛋白質是必不可少的,而ATP水解所產生的能量主要是用於蛋白質的去摺疊、核心顆粒的孔道開放 還是兩者皆有,則還不清楚。截止到2006年,26S蛋白酶體的結構還沒有獲得解析。
19S顆粒的每個組分都有它們自己的調控作用。一個近期鑒定出的癌蛋白Gankyrin(英語:Gankyrin)是19S顆粒的組分之一,可以與細胞周期蛋白依賴性激酶CDK4緊密結合,並且通過與泛素連接酶MDM2(英語:MDM2)的結合,在識別泛素化的P53蛋白中發揮作用。Gankyrin具有抗凋亡作用,其被發現在一些類型的腫瘤細胞(如肝癌細胞)中過表達。
20S核心顆粒也可以與第二種調節顆粒,即11S顆粒相結合。11S調節顆粒又被稱為PA28或REG。它是七聚體結構,不包含任何ATP酶,能夠促進
短肽而不是完整的蛋白質的降解。這可能是因為由11S顆粒與核心顆粒所組成的複合物無法將大的底物去摺疊。11S顆粒的調控機制與19S顆粒的機制類似,是通過其亞基的C末端結合核心顆粒,並誘發α環發生構象變化,從而打開20S核心顆粒的“門”,使得底物蛋白質可以進入核心顆粒。 11S顆粒的表達受γ干擾素的誘導,並且負責與免疫蛋白酶體的β亞基一起生成結合到主要組織相容性複合體上的肽段。
11S和19S調節顆粒都是多亞基的複合物,而實際上真核生物中還存在著以單個蛋白結合20S顆粒的調節蛋白(酵母中)。PA200的分子量高達200kDa,其主要定位於
細胞核中,可以直接結合併激活20S顆粒。 PA200可能參與了DNA雙鏈斷裂的修復。
需要被蛋白酶體降解的蛋白質會先被連接上泛素作為標記,即蛋白質上的一個賴氨酸與泛素之間形成共價連接。這一過程是一個三酶級聯反應,即需要有由三個酶催化的一系列反應的發生,整個過程被稱為泛素化信號通路。在第一步反應中,泛素活化酶(又被稱為E1)水解ATP並將一個泛素分子腺苷酸化。接著,泛素被轉移到E1的活性中心的
半胱氨酸殘基上,並伴隨著第二個泛素分子的腺苷酸化。被
腺苷酸化的泛素分子接著被轉移到第二個酶,泛素交聯酶(E2)的半胱氨酸殘基上。最後,高度保守的泛素連接酶(E3)家族中的一員(根據底物蛋白質的不同而不同)識別特定的需要被泛素化的靶蛋白,並催化泛素分子從E2上轉移到靶蛋白上。靶蛋白在被蛋白酶體識別之前,必須被標記上至少四個泛素單體分子(以多泛素鏈的形式)。因此,是E3使得這一系統具有了底物特異性。 蛋白的數量依賴於生物體和細胞類型,人體中就存在大量不同的E3蛋白,這說明泛素-蛋白酶體系統可以作用於數量巨大的靶蛋白。
多泛素化后的蛋白質是如何被蛋白酶體所識別的,還沒有完全弄清。泛素受體蛋白的N末端具有一個類泛素結構域,以及一至多個泛素結合結構域。類泛素結構域可以被19S調節顆粒所識別,而泛素結合結構域可以通過形成三螺旋束來結合泛素。這些受體蛋白可能能夠結合多泛素化的蛋白質並將其攜帶到蛋白酶體,而關於這種結合的特異性和調控機制還不清楚。但最近有研究者發現,調節顆粒上的亞基可以發揮泛素受體的功能。
泛素蛋白自身由76個殘基所組成,以“泛素”為名是因為它在生物體中廣泛存在:具有高度保守的序列並且存在於所有已知的真核生物體中。真核生物中編碼泛素的基因以串聯重複(英語:tandem repeat)的方式排列,這可能是因為大量
轉錄的需要,為細胞生產足夠多的泛素。有人提出泛素是目前發現的進化速度最慢的蛋白質。
泛素化信號通路。其中,“Ub”表示泛素。
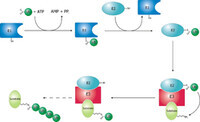
泛素化信號通路。其中,“Ub”表示泛素。
泛素化后的蛋白質(以下稱為底物蛋白)被19S調節顆粒所識別,這一過程是一個ATP依賴的結合過程。然後,底物蛋白必須進入20S核心顆粒的內部孔道,以便與位於其中的水解活性位點接觸。由於20S顆粒的孔道相對狹窄,而且兩端由α環中亞基的N末端控制開關,所以底物蛋白在進入核心顆粒之前必須至少部分去摺疊。將去摺疊的蛋白質傳遞進入核心顆粒的過程被稱為“移位”(translocation),而移位必須發生在去泛素化之後。但目前對於底物蛋白的去泛素化和去摺疊機制還不了解。在整個降解反應過程中,那一步是限速步取決於底物蛋白的類別;對於一些蛋白質,去摺疊過程是限速步,而對於另一些蛋白質,可能是去泛素化為限速因子。至於哪些底物蛋白在移位之前必須去摺疊,還未有結論,而牢固的三級結構和一些特殊的非局部相互作用,如二硫鍵,能夠抑制降解。
由α亞基所形成的“門”可以阻止長於四個殘基的多肽進入20S顆粒的內部。在識別步驟開始前結合上的ATP分子在移位發生前被水解,而對於水解產生的能量是用於蛋白質去摺疊 還是“門”的打開 還有爭議。26S蛋白酶體在存在無法水解的ATP類似物(即無法獲得水解產生的能量)的情況下,依然可以降解去摺疊的蛋白質,但卻無法降解摺疊的蛋白質;這一結果說明ATP水解所產生的能量至少部分被用於蛋白質去摺疊。在19S帽子處於ATP結合狀態時,去摺疊的底物蛋白可以由促進擴散作用,傳遞通過開啟的“門”。
球蛋白去摺疊的機制是基本類似的,但在一定程度上也取決於蛋白質的氨基酸序列。研究者發現含有較長的
甘氨酸或
丙氨酸序列可以抑制去摺疊,從而降低蛋白酶體的降解效率;其結果是生成含有部分去摺疊蛋白質的混合物,這可能是由於ATP水解和去摺疊步驟之間的脫節所導致的。自然界中的一些蛋白質也有這樣的甘氨酸-丙氨酸重複序列存在,如蠶絲中的絲心蛋白(英語:fibroin);值得一提的是,特定的人類皰疹病毒基因的表達產物也含有這樣的序列,通過抑制蛋白酶體的作用,阻止了抗原呈遞到主要組織相容性複合體上,從而有助於病毒的繁殖。
20S核心顆粒的一個剖面圖,顯示了活性位點的位置。其中,α亞基用綠色的球來表示,β亞基的蛋白骨架顯示為飄帶,並且不同的
多肽鏈用不同的顏色表示。小的粉色球表示每個亞基的活性位點中蘇氨酸殘基的位置。淡藍色的化學結構為結合在活性位點上的抑製劑硼替佐米。
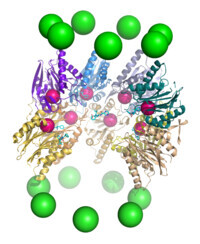
20S核心顆粒的一個剖面圖,顯示了活性位點的位置。其中,α亞基用綠色的球來表示,β亞基的蛋白骨架顯示為飄帶,並且不同的多肽鏈用不同的顏色表示。小的粉色球表示每個亞基的活性位點中蘇氨酸殘基的位置。淡藍色
蛋白質的降解由20S核心顆粒中的β亞基進行,其機制被認為是蘇氨酸依賴的親核攻擊。這一機制可能需要有一個結合的水分子參與活性的蘇氨酸上羥基的去質子化。降解發生在核心顆粒中間的兩個β環內的孔道里,一般不生成部分降解的產物,而是將底物蛋白完全降解為長度一定的肽段;肽段的長度一般為個殘基,但根據生物體和底物蛋白的不同,長度範圍可以從個殘基不等。決定分解產物中肽段長度的機制,目前還沒有完全弄清。雖然具有催化活性的三個β亞基具有共同的降解機制,但它們對於底物的特異性卻略有不同,分別為類胰凝乳蛋白酶型、類胰蛋白酶型和肽谷氨醯基肽水解型。這種對於底物特異性的差異是來自於靠近活性位點的局部殘基與底物之間的相互作用的不同。每一個具有催化活性的β亞基也都含有一個降解所必需的保守的賴氨酸。
雖然蛋白酶體通常生成非常短的降解片斷,但在一些情況下,這些降解產物自身是具有生物學活性的功能分子。特定的
轉錄因子,包括哺乳動物的(英語:複合物中的一個組分,合成后是以無活性的前體分子存在,在經過泛素化和蛋白酶降解后,才轉變為活性分子。這種降解需要蛋白酶體剪切蛋白質的中間部分,而不是通常情況下的從蛋白質的一端開始的剪切。有人提出,需要被剪切的中間部分為一個長的環(英語:loop (biochemistry)),位於蛋白表面,從而可以作為蛋白酶體的底物進入其內部孔道,而蛋白質的其他部分依然在孔道外,並不會被降解。在
酵母蛋白中也發現了類似的現象;這種選擇性降解被稱為“受調控的泛素-蛋白酶體依賴的剪切”(regulated ubiquitin/proteasome dependent processing)。
雖然大多數的蛋白酶體的底物必須在降解之前被泛素化,但仍然有一些例外的情況,尤其是在蛋白酶體參與蛋白質的翻譯后處理過程中。一個主要的例子是蛋白酶體通過將p105(英語:p105)蛋白剪切為p50(英語:P50 (biochemistry))蛋白來激活。一些由於存在無結構區域(英語:intrinsically unstructured proteins)而被推測具有不穩定性的蛋白質也可以通過非泛素依賴的途徑被降解。鳥氨酸脫羧酶是最著名的非泛素依賴途徑中蛋白酶體的底物。對於關鍵的細胞周期調控因子,如P53蛋白的非泛素依賴的降解機制已經有報道,雖然p53蛋白也可以通過泛素依賴的途徑被降解。此外,在一定的細胞應激條件下,結構不正常、錯誤摺疊或者過度氧化的蛋白質也都會進入非泛素依賴的和非19S顆粒依賴的降解途徑。

大腸桿菌中的hslV
中的hslV(英語:HslV)(藍色)和hslU(英語:HslU)複合物(紅色)。這一由熱休克蛋白組成的複合物被認為類似於現在蛋白酶體的祖先。
20S蛋白酶體在真核生物中廣泛存在且必不可少。一些原核生物,包括許多古菌和細菌中的
放線菌也含有20S蛋白酶體的同源體,即大多數細菌都含有的熱休克基因hslV(英語:HslV)和hslU(英語:hslU),這兩個基因所編碼的蛋白質可以形成雙層環狀多聚體和ATP酶。一些研究者認為HslV蛋白很可能類似於20S蛋白酶體的祖先。一般來說,HslV蛋白對於細菌不是必要的,且並非所有的細菌都含有這一蛋白,而
原生生物同時含有20S蛋白酶體和HslV蛋白系統。
序列分析顯示,催化性的β亞基在進化過程中分化得比結構性的α亞基要早。表達20S蛋白酶體的細菌中,其β亞基與古菌以及真核生物的β亞基具有高度的序列相似性,而α亞基的序列相似程度則低得多。細菌中存在20S蛋白酶體可能是基因水平轉移的結果,而真核生物中各亞基的分化則應是多次基因重複的結果。
細胞周期進程是由一系列細胞周期蛋白依賴性激酶(CDK)來進行調控的,而CDK則是由細胞周期蛋白(cyclin)來激活。有絲分裂的細胞周期蛋白,在細胞中只有幾分鐘壽命,是所有已知的細胞內蛋白中壽命最短的。在CDK-cyclin複合物行使了它的功能之後,複合物中的cyclin就會被多泛素化並由蛋白酶體降解,從而保證了細胞周期的正常運轉。尤其是在細胞退出有絲分裂期時,作為調控組分的周期蛋白B(英語:cyclin B)需要從有絲分裂促進因子上脫落下來,而這一解離過程依賴於蛋白酶體的參與。
細胞周期檢控點,如G1期和S期之間的后限制點檢查,也需要蛋白酶體降解周期蛋白A(英語:cyclin A),而cyclin A的泛素化由一個名為後期促進複合物(anaphase promoting complex,APC)的E3泛素連接酶來進行。 APC蛋白和蛋白複合物(即SCF複合物)是降解cyclin和控制檢控點的兩個關鍵調控因子;SCF複合物自身則由APC蛋白來調控,由於Skp2蛋白(SCF複合物中的轉接蛋白)可以在G1期到S期的過渡期中抑制SCF複合物的活性,因此通過泛素化Skp2蛋白,APC蛋白就可以激活SCF複合物。
[編輯]調控植物生長
在植物中,
生長素或
植物激素的作用是調控植物生長的方向和向性,它們通過細胞信號通路來誘導一系列轉錄因子抑制蛋白(蛋白)進入蛋白酶體降解途徑。這些抑制蛋白由SCFTIR1蛋白或者由與auxin受體蛋白TIR1結合的SCF蛋白進行泛素化。蛋白降解后,對auxin反應因子(ARF)家族的轉錄因子的抑制就被解除,從而誘導ARF基因的表達。 ARF被激活所導致的結果因植物類型和發育水平的不同而有所差異,但都參與了對根和葉脈生長的指導。ARF蛋白和蛋白之間配對的特異性被認為是ARF的去抑制作用具有反應特異性的原因。
細胞內外的信號都能夠誘導細胞凋亡或編程性細胞死亡。其結果是細胞內部的組分發生解構,這主要是由特定的蛋白酶半胱天冬酶來完成,但同時蛋白酶體也可能在細胞凋亡過程中扮演了多種重要角色。蛋白酶體參與細胞凋亡進程的推測是基於凋亡發生前,細胞中泛素化蛋白質以及在數量上的增加這一現象;並且,在細胞凋亡過程中,原本定位於細胞核的蛋白酶體被發現能夠移位到調亡小泡的外膜。
蛋白酶體的抑制作用可以影響不同類型細胞的調亡誘導,在大多數已被研究的細胞類型中,抑制蛋白酶體可以促進細胞調亡。但一般而言,蛋白酶體並非是細胞調亡所必需的因子。而且,對於一些細胞系,特別是原代培養的靜止和分化的細胞,如胸腺細胞和
神經元細胞,暴露於蛋白酶體抑製劑反而阻止了細胞的凋亡。這一作用機制目前還不清楚,但有人推測這種現象只特異性地發生於靜止狀態的細胞或者這是由於促細胞凋亡激酶JNK(英語:JNK)的活性差異所導致的。由於
蛋白酶抑製劑可以誘發處於快速分裂中的細胞(如癌細胞)的凋亡,因此一些蛋白酶抑製劑已經被開發並作為化療藥品被用於治療癌症。
蛋白酶體直接參與了適應性免疫系統的運作,並在其中扮演著關鍵角色。肽類抗原是由主要組織相容性複合物(MHC)類型I蛋白傳遞到抗原呈遞細胞表面。這些肽段是來自被蛋白酶體降解的侵入機體的病原體。雖然一般的蛋白酶體就可以參與這一進程,但實際上起主要作用的是一種特殊的複合物,其可以生成合適大小和成分的降解片斷以供
MHC結合。這種複合物的組成蛋白的表達是由γ干擾素所誘導;當免疫反應發生時,這些蛋白質,包括11S調節顆粒(主要作用為調節MHC的結合肽段的產生)和特殊的β亞基(,具有不同的底物特異性)的表達就會增加。這種由特殊的β亞基參與形成的複合物就被稱為“免疫蛋白酶體”。另一種有所變化的β5亞基,β5t,在
胸腺中表達,能夠形成胸腺獨有的“胸腺蛋白酶體”("thymoproteasome"),參與
T細胞的發育調控。
MHC類型I蛋白的配基結合強度取決於配基C末端的組成,因為肽段配基是通過氫鍵和與MHC表面的"B pocket"近接觸來結合的。許多MHC類型I蛋白趨向於結合疏水性殘基,而免疫蛋白酶體複合物就可以更多地生成具有疏水性C末端的肽段。
由於蛋白酶體參與生成活性形勢的(英語:)(一種抗凋亡和促炎症調控因子,調控細胞因子的表達),因此,蛋白酶體被認為與炎症反應和
自身免疫性疾病相關。蛋白酶體活性水平的提高與包括紅斑性狼瘡和
類風濕性關節炎在內的自身免疫性疾病相關。
蛋白酶體抑製劑硼替佐米的化學結構。採用硼替佐米的化學療法可以有效地治療
多發性骨髓瘤。
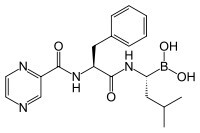
蛋白酶體抑製劑硼替佐米的化學結構。採用硼替佐米的化學療法可以有效地治療多發性骨髓瘤。
硼替佐米結合於酵母蛋白酶體的核心顆粒上。硼替佐米分子位於圖的正中;其中,粉色表示
碳原子,藍色表示氮原子,紅色表示
氧原子,黃色表示硼原子。環繞在硼替佐米分子周圍的是蛋白質局部表面,其中藍色的部分是發揮催化作用的蘇氨酸殘基,由於結合上硼替佐米,因而失去了催化活性。
蛋白酶體抑製劑對於人工培養的細胞具有有效的抗腫瘤活性,通過降解受調控的促生長細胞周期蛋白來誘導腫瘤細胞的凋亡。這種可以選擇性地誘導腫瘤細胞凋亡的方法被證明在動物模型以及人體試驗中都非常有效。硼替佐米是第一種用作化學治療藥物的蛋白酶體抑製劑,由千年製藥公司(英語:Millennium Pharmaceuticals)開發,市場名稱為
Velcade。硼替佐米主要被用於多發性骨髓瘤的治療 值得一提的是,多發性骨髓瘤會導致血清中蛋白酶體水平的提高,而成功的化療可以將蛋白酶體的水平恢復到正常範圍。動物研究顯示硼替佐米可能對死亡率極高的
胰腺癌也有顯著的臨床效果。對於硼替佐米在治療
B細胞相關癌症的臨床前和早期臨床研究已經開始,特別是一些類型的非霍奇金氏淋巴瘤。
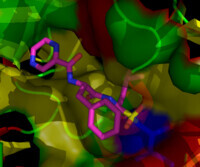
硼替佐米結合於酵母蛋白酶體的核心顆粒上。硼替佐米分子位於圖的正中;其中,粉色表示碳原子,藍色表示氮原子,紅色表示氧原子,黃色表示硼原子。環繞在硼替佐米分子周圍的是蛋白質局部表面,其中藍色的部分是發揮催
(英語:ritonavir),市場名稱為Norvir,是用於治療
艾滋病的一種蛋白酶抑製劑。近期的研究發現,利托那韋不僅可以抑制蛋白酶,對蛋白酶體也有抑制作用,特別是對類
胰凝乳蛋白酶型的蛋白酶體,但對類
胰蛋白酶的蛋白酶體則有部分的促進作用。對於動物模型的研究表明利托那韋可能對神經膠質瘤細胞的生長有抑制作用。
將蛋白酶體抑製劑應用於
自體免疫性疾病也大有前景。目前這一方面的研究主要是利用動物模型來進行。例如,在對植有人類皮膚的小鼠的研究中發現,用蛋白酶體抑製劑處理過後,原本因患牛皮癬而受損的皮膚所有減少;在一些
嚙齒動物模型中發現,蛋白酶體抑製劑對於
哮喘也有一定的治療作用。
蛋白酶體的標記和抑制作用也被用於在實驗室中針對細胞中蛋白酶體活性所進行的“體外”(in vitro)和“體內”(in vivo)研究。實驗室中最常用的抑製劑為乳胞素(英語:lactacystin),一種由
鏈黴菌合成的天然產物。帶有熒光基團的抑製劑也已經被開發應用於特異性標記組裝好的蛋白酶體上的活性位點。