共找到2條詞條名為氣孔的結果 展開
- 植物器官上皮許多小開孔之一
- 鑄造缺陷之一
氣孔
植物器官上皮許多小開孔之一
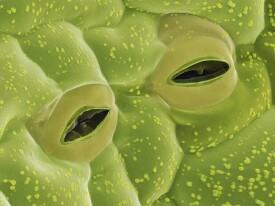
氣孔,葉、莖及其他植物器官上皮上許多小的開孔之一,是植物表皮所特有的結構。氣孔通常多存在於植物體的地上部分,尤其是在葉表皮上,在幼莖、花瓣上也可見到,但多數沉水植物則沒有。
狹義上常把保衛細胞之間形成的凸透鏡狀的小孔稱為氣孔。保衛細胞區別於表皮細胞是結構中含有葉綠體,只是體積較小,數目也較少,片層結構發育不良,但能進行光合作用合成糖類物質。有時也伴有與保衛細胞相鄰的2—4個副衛細胞。把這些細胞包括在內是廣義的氣孔(或氣孔器)。緊接氣孔下面有寬的細胞間隙(氣室)。氣孔在碳同化、呼吸、蒸騰作用等氣體代謝中,成為空氣和水蒸氣的通路,其通過量是由保衛細胞的開閉作用來調節,在生理上具有重要的意義。
以裸子植物為中心對氣孔的形成過程和親緣關係十分重視。氣孔是從原表皮細胞中發生的,氣孔母細胞(stomatal mother cell)橫分裂為三,中央細胞再分為二,成為保衛細胞,左右二細胞則成為副衛細胞的形式[復唇型(syndetocheilie type),相反,也有母細胞僅二分為保衛細胞的形式[單唇形(haplocheilic type) ,後者被視為原始型。這兩種形式在裸子植物系統分類上受到重視(R.Flor-in,1951),單唇型在蘇鐵蕨類(Cycadofilices)、蘇鐵類(Cycadinae)、苛得狄類(Cordaitinae)、銀杏類(Ginkgoinae)、針葉樹類(Coniferae)、麻黃類(Ephedrales)(狹義)可見到,復唇型則在本納蘇鐵類(Bennettitales)、百歲蘭類(Welwitschiales)、買麻藤類(Gnetales)可見到。
![氣孔[植物器官上皮許多小開孔之一]](https://i1.twwiki.net/cover/w200/m5/c/m5cd1f6e0cf906f991e74f77bdfecaf90.jpg)
氣孔[植物器官上皮許多小開孔之一]
唇形科(Labiafae),報春花科(Primulaceae)其他很多濕地植物]和下陷的報春花[針葉樹類(松柏類)、木賊科(Eguisetaceae)、仙人掌科(Cact-aceae)、夾竹桃(Nerium indicum)]等,均具有生態學方面的重要意義。把角苔(Anthoceros)擬為高等植物的原始型,就是由於重視其孢子體上有氣孔。
總的來講,不同植物的葉、同一植物不同的葉、同一片葉的不同部位(包括上、下表皮)都有差異,且受客觀生境條件的影響。浮水植物只在上表皮分佈,陸生植物葉片的上下表皮都可能有分佈,一般陽生植物葉下表皮較多,上表皮接受陽光,水分散失快,所以上表皮少。
![氣孔[植物器官上皮許多小開孔之一]](https://i1.twwiki.net/cover/w200/m9/1/m91ad9dbb189e7aeaf244744233c93376.jpg)
氣孔[植物器官上皮許多小開孔之一]
無規則型
保衛細胞周圍無特殊形態分化的副衛細胞;
不等型
保衛細胞周圍有三個副衛細胞圍繞;
平行型
在保衛細胞的外側面有幾個副衛細胞與其長軸平行;
橫列型
一對副衛細胞共同與保衛細胞的長軸成直角。圍成氣孔間隙的保衛細胞形態上也有差異,大多數植物的保衛細胞呈腎形,近氣孔間隙的壁厚,背氣孔間隙的壁薄;稻、麥等植物的保衛細胞呈啞鈴形,中間部分的壁厚,兩頭的壁薄。
氣孔的開關與保衛細胞的水勢有關,保衛細胞水勢下降而吸水膨脹,氣孔就張開,水勢上升而失水縮小,使氣孔關閉。
引起保衛細胞水勢的下降與上升的原因主要存在以下學說。
澱粉-糖轉化學說
(starch-sugar conversion theory)
很早以前已觀察到,pH影響磷酸化酶反應(在pH6.1~7.3時,促進澱粉水解;在pH2.9~6.1時,促進澱粉合成):
澱粉-糖轉化學說認為,植物在光下,保衛細胞的葉綠體進行光合作用,導致CO2濃度的下降,引起pH升高(約由5變為7),澱粉磷酸化酶促使澱粉轉化為葡萄糖-1-P,細胞里葡萄糖濃度高,水勢下降,副衛細胞(或周圍表皮細胞)的水分通過滲透作用進入保衛細胞,氣孔便開放。黑暗時,光合作用停止,由於呼吸積累CO2和H2CO3,使pH降低,澱粉磷酸化酶促使糖轉化為澱粉,保衛細胞里葡萄糖濃度低,於是水勢升高,水分從保衛細胞排出,氣孔關閉。試驗證明,葉片浮在pH值高的溶液中,可引起氣孔張開;反之,則引起氣孔關閉。
但是,事實上保衛細胞中澱粉與糖的轉化是相當緩慢的,因而難以解釋氣孔的快速開閉。試驗表明,早上氣孔剛開放時,澱粉明顯消失而葡萄糖並沒有相應增多;傍晚,氣孔關閉后,澱粉確實重新增多,但葡萄糖含量也相當高。另外,有的植物(如蔥)保衛細胞中沒有澱粉。因此,用澱粉-糖轉化學說解釋氣孔的開關在某些方面未能令人信服。
無機離子吸收學說
(inorganic ion uptake theory)
該學說認為,保衛細胞的滲透勢是由鉀離子濃度調節的。光合作用產生的ATP,供給保衛細胞鉀氫離子交換泵做功,使鉀離子進入保衛細胞,於是保衛細胞水勢下降,氣孔就張開。1967年日本的M.Fujino觀察到,在照光時漂浮於KCl溶液表面的鴨跖草保衛細胞鉀離子濃度顯著增加,氣孔也就開放;轉入黑暗或在光下改用Na、Li時,氣孔就關閉。撕一片鴨跖草表皮浮於KCl溶液中,加入ATP就能使氣孔在光下加速開放,說明鉀離子泵被ATP開動。用電子探針微量分析儀測量證明,鉀離子在開放或關閉的氣孔中流動,可以充分說明,氣孔的開關與鉀離子濃度有關。
蘋果酸生成學說
(malate production theory)
人們認為,蘋果酸代謝影響著氣孔的開閉。在光下,保衛細胞進行光合作用,由澱粉轉化的葡萄糖通過糖酵解作用,轉化為磷酸烯醇式丙酮酸(PEP),同時保衛細胞的CO2濃度減少,pH上升,剩下的CO2大部分轉變成碳酸氫鹽(HCO3),在PEP羧化酶作用下,HCO3與PEP結合,形成草醯乙酸,再還原為蘋果酸。蘋果酸會產生H+,ATP使H-K交換泵開動,質子進入副衛細胞或表皮細胞,而K進入保衛細胞,於是保衛細胞水勢下降,氣孔就張開。
此外,氣孔的開閉與脫落酸(ABA)有關。當將極低濃度的ABA施於葉片時,氣孔就關閉。後來發現,當葉片缺水時,葉組織中ABA濃度升高,隨後氣孔關閉。
光照引起的氣孔運動
保衛細胞的葉綠體在光照下進行光合作用,利用CO2,使細胞內pH值增高,澱粉磷酸化酶水解澱粉為磷酸葡萄糖,細胞內水勢下降.保衛細胞吸水膨脹,氣孔張開;黑暗裡呼吸產生的CO2使保衛細胞的pH值下降,澱粉磷酸化酶又把葡萄糖合成為澱粉,細胞液濃度下降,水勢升高,保衛細胞失水,氣孔關閉。保衛細胞的滲透系統也可由K 來調節。光合作用光反應(環式與非環式光合磷酸化)產生ATP,通過主動運輸逆著離子濃度差吸收K ,降低保衛細胞水勢,吸水使氣孔張開。注意:①如果光照強度在光補償點以下,氣孔關閉;②在引起氣孔張開的光質上以紅光與藍紫光效果最好;③景天科植物夜晚氣孔張開,吸收和貯備CO2(形成蘋果酸貯於液泡中),白天氣孔關閉,蘋果酸分解成丙酮酸釋放CO2進行光合作用。
二氧化碳影響氣孔運動
低濃度CO2促進氣孔張開,高濃度CO2使氣孔迅速關閉,無論光照或黑暗皆如此。抑制機理可能是保衛細胞pH下降,水勢上升,保衛細胞失水,必須在光照一段時間待CO2逐漸被消耗后,氣孔才迅速張開。
溫度影響氣孔運動
氣孔張開度一般隨溫度的上升而增大,在30℃左右達到最大,低溫(如10℃以下)雖長時間光照,氣孔仍不能很好張開,主要是澱粉磷酸化酶活性不高之故,溫度過高會導致蒸騰作用過強,保衛細胞失水而氣孔關閉。
葉片含水量影響氣孔運動
白天若蒸騰過於強烈,保衛細胞失水氣孔關閉,陰雨天葉子吸水飽和,表皮細胞含水量高,擠壓保衛細胞,故白天氣孔也關閉。
風
微風時對氣孔的打開有促進作用,因為微風可以適當降低葉片周圍的濕度。大風則促使氣孔關閉。
化學物質
醋酸苯汞、阿特拉津(2-氯-4-乙氨基-6-異丙氨基均三氮苯)、乙醯水楊酸等能抑制氣孔開放,降低蒸騰。脫落酸的低濃度溶液灑在葉表面,可抑制氣孔開放達數天,並且作用快,在2~10分鐘內可使多種植物氣孔開始關閉。細胞分裂素可促進氣孔開放。
[blowhole]∶金屬鑄錠和鑄件中的孔
鑄件氣孔的分類,主要有三類:侵入性氣孔、析出性氣孔、反應性氣孔。主要是由於金屬溶液中含有過多的氣體或者金屬溶液中發生反應生成氣體無法有效的排出而生成。
[air hole]∶接受或放出空氣的孔
基本信息
- 中文名
- 氣孔
- 外文名
- stomata
- 特點
- 植物表皮所特有的結構
- 性質
- 植物器官上皮許多小開孔之一
- 釋義
- 葉、莖及其他植物器官上皮上許多小的開孔之一
- 存在位置
- 植物體的地上部分
- 作用
- 在碳同化、呼吸、蒸騰作用等氣體代謝中,成為空氣和水蒸氣的通路