細胞學
在光學顯微鏡水平上研究細胞的學科
在光學顯微鏡水平上研究細胞的化學組成、形態、結構及功能的學科。研究細胞結構和功能的生物學分支學科。細胞是組成有機體的形態和功能的基本單位,自身又是由許多部分構成的。關於結構的研究不僅要知道它是由哪些部分構成的,而且要進一步搞清每個部分的組成。相應地,關於功能不僅要知道細胞作為一個整體的功能,而且要了解各個部分在功能上的相互關係。
細胞學是研究細胞的形態、結構和功能以及與細胞生長、分化、進化等相關聯的生物學的一個分支學科。生物體的生理功能及一切生命現象,都是以細胞為基本單位而表達的。因此,不論對生物體的遺傳、發育以及生理機能的了解,還是對於作為醫療基礎的病理學、藥理學等以及農業的育種等,細胞學都至關重要。
任何生物現象無不來自細胞的功能,所以生物學的所有領域都與細胞學有關。細胞學獨立為一門學科是從確立M.J.Schleiden(1838)和T.Schwann(1839)的細胞學說開始的。隨著生物組織培養、顯微解剖、電子顯微鏡、紫外線顯微鏡、相差顯微鏡、超速離心分離法以及冷凍乾燥法等技術的發展,對細胞結構、有絲分裂以及細胞內滲透壓和細胞膜透性等細胞生理功能方面的理論得到發展和證實。進入20世紀之後,細胞學的應用越來越引起重視。
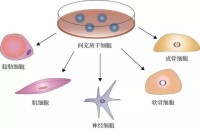
細胞學
對於研究細胞起了巨大推動作用的是M.J.施萊登和T.A.H.施萬。前者在1838年描述了細胞是在一種粘液狀的母質中經過一種像是結晶樣的過程產生的,而且首先產生出核(還發現核仁)。他並且把植物看作細胞的共同體,就好像水螅蟲的群體一樣。在他的啟發下施萬堅信動、植物都是由細胞構成的。他積累了大量事實,指出二者在結構和生長中的一致性,於1839年提出了細胞學說。與此同時,捷克動物生理學家J.E.浦肯野提出原生質的概念;德國動物學家C.T.E.von西博爾德(1845)斷定原生動物都是單細胞的。德國病理學家R.C.菲爾肖(1855)在研究結締組織的基礎上提出“一切細胞來自細胞”的名言,並且創立了細胞病理學。德國動物學家M.舒爾策在1861年對細胞下了定義:“細胞是一團具有一切生命特徵的原生質,細胞核處於其中。”
從19世紀中期到20世紀初,關於細胞核的研究,有了長足的進展。
1、1875年,德國植物學家E.A.施特拉斯布格首先敘述了植物細胞中的著色物體而且斷定同種植物各自有一定數目的著色物體。
2、1880年,巴拉涅茨基描述了著色物體的螺旋狀結構,翌年普菲茨納發現了染色粒。
3、1885年,德國學者C.拉布爾提出著色物體數目恆定的規律。
4、1888年,W.瓦爾代爾把核中的著色物體正式命名為染色體。
5、1891年,德國學者H.亨金在昆蟲的精細胞中觀察到X染色體。
6、1902年,W.L.史蒂文斯、E.B.威爾遜等發現了Y染色體。
細胞分裂現象,在此期間已經受到重視,並進行了仔細分析。
1、1867年,德國植物學家W.霍夫邁斯特在植物,A.施奈德1873年在動物,分別比較詳細地敘述了間接分裂。
2、1882年,德國細胞學家W.弗勒明在發現了染色體的縱分裂之後提出了有絲分裂這一名詞以代替間接分裂,E.霍伊澤爾描述了在間接分裂時的染色體分佈;在他之後,E.A.施特拉斯布格把有絲分裂劃分為:前期、中期、後期、末期;他和其他學者還在植物中觀察到減數分裂,經過進一步研究終於區別出單倍體和雙倍體染色體數目。
3、1933年,H.鮑爾在蚊子的馬爾皮基氏管細胞中發現了多線染色體。
4、1934年,T.S.佩因特在果蠅,R.L.金和H.W.比姆斯在搖蚊中,也發現這種構造。
多線染色體是一種存在於雙翅目幼蟲的某些腺體細胞中的巨大染色體,在果蠅中其長度大約是正常染色體的100倍,每條染色體由許多條(可多到400條)染色纖維組成,在整條染色體上顯示染色深的帶區和染色淺的間帶區。它的形成是由於核內有絲分裂(只有染色體分裂而核不分裂),因而每條多線染色體實際上是由許多染色體形成的。這種染色體體積龐大,有利於對染色體的精細構造進行分析。此外,還可根據多線染色體上的脹泡判斷其功能活動的情況。
直到70年代,在電子顯微鏡下觀察到核小體;此後不久,結合生化提取,觀察到分裂中期的染色體是以所謂的支架蛋白為核心,DNA纖維由此環狀地向四周伸展出去形成螺旋化。
對細胞質的認識落後於對細胞核或染色體的認識,而且經歷很長時期才得到改善。
1、1865年,C.弗羅曼認為細胞中含有纖維狀物質交織成框架或網狀。
2、1875年,德國生物學家O.赫特維希發現了中心體。
3、1882年,W.弗勒明錯誤地把所看到的線粒體、紡錘絲以及固定樣品中的其他纖維狀構造推而廣之,認為細胞質是由埋藏在基質中的這些絲狀成分構成的。
4、1886年,德國組織學家R.阿爾特曼甚至認為一定的小顆粒是最簡單的、活的、“細胞的基本有機體”,由於它們的特殊方式的集聚而構成細胞;這可能也是由於誤認了線粒體以及分泌和貯藏顆粒。
5、1888年,德國動物學家O.比奇利提出了蜂窩或泡沫學說,即:細胞質是由較粘的物質(透明質hyalopla-sm)形成的精細的蜂窩狀構造構成的,其中充滿另一種稱之為細胞液(enchylema)的物質。這個學說在一定程度上符合實際情況,也比較容易被人接受,因為比奇利不是根據對固定的標本觀查,而是根據對原生動物的活體觀察提出的。(註:原生動物太陽蟲的細胞質確實是泡沫狀的──關於原生動物是否單細胞的問題爭論了差不多半個世紀,直到1875年經比奇利研究纖毛蟲后才予以肯定──因此泡沫狀學說維持的時間最長。)
6、1895年,高爾基發現了被他稱之為Apparato reticulare interno的網狀結構物質(后稱:高爾基器)。
7、1897年,C.本達發現了線粒體並命名,對於它的存在意見比較一致。在一些細胞中經一定的固定劑固定后,可被一定的染料染色,也可在活體中觀察到。但是在光學顯微鏡下其形狀各式各樣,或是線狀或是顆粒狀或是一串顆粒;至於是否存在於動物的各種細胞內或一切生物體的細胞內,當時還沒有定論。
8、1899年,加尼耶在研究各類腺體細胞時發現細胞質中含有嗜鹼性的呈現動態變化的絲狀或棒狀的結構,認為這不是細胞質的內含物,而是細胞質的組成部分,因而命名為動質(后稱內質網),並且對此做了詳細的敘述。
進入20世紀之後,尤其是電子顯微鏡得到廣泛使用,標本的包埋、切片一套技術逐漸完善,才有了很大改變。通過大量的工作,不僅弄清楚了從前在光學顯微鏡下可以看到而又看不清,或者尚有爭議的細胞器,如線粒體、高爾基器、中心體、內質網、纖毛、鞭毛等構造,而且還發現了許多從前未曾看到過的構造如溶酶體、過氧化酶體、核糖體以及構成細胞骨架的各種纖維物質。
20世紀40年代后,利用高壓電鏡觀察到了由1~10埃粗細的纖維組成的支撐著各種細胞器的微梁系統,而且看到了細胞的各種膜。在電鏡下斷定了所有的膜都是75~100埃厚的三層結構(稱之為單位膜)。不僅如此,一個細胞的各部分膜都是相連的,質膜與內質網,內質網與高爾基器或核膜相連。核膜是雙層的,由內外兩層膜構成,並且具有有一定結構的核膜孔,通過它,細胞質的物質和細胞核的物質得以交流。在質膜上還發現了細胞間連結:橋粒、緊密連接和間隙連接等。這些結構與細胞間的結合或細胞間的物質交流有關;利用冰凍蝕刻技術,可以更好地觀察它們。
細胞學的研究,在相當程度上受到其他學科的推動,根據各學科的影響大致地可以劃分幾個階段,當然這些階段不可能截然分開。
胚胎學的影響。對細胞功能,不能像研究結構那樣,在一團組織里找一個細胞作為研究對象。卵子是一個細胞,在無法得到單個的細胞進行研究的年代,利用它是極為方便的材料。既然用卵子,研究它各部分的作用當然要根據對發育中的影響來判斷。這涉及胚胎學問題。但是如果用雜交研究異種精核的功能,則需要根據異種性狀的出現來判斷,這就涉及到遺傳的問題。早期在這方面的工作基本上是由胚胎學家進行的,其特點是綜合性的研究,不是單純地從細胞的角度研究卵子,而是拿卵子作為細胞來研究與發育、遺傳等有關的問題。一些重大的問題都已勾劃出來,因而在學術思想上對以後有深刻的影響。O.赫特維希和R.von赫特維希弟兄1887年用海膽作材料,首先看到活的卵子的受精,並且對受精進行了實驗分析。如果分別地考慮細胞質和細胞核在發育中的作用,則T.H.博韋里對在馬蛔蟲中發現的染色質消減的現象的分析,證明影響消減的因素存在於細胞質中。此外,對卵裂球予以編號,以追蹤每一裂球的來龍去脈的細胞譜系工作,關於卵黃含量不同的各種卵子其卵裂類型的研究,都指出卵子中細胞質的分佈,影響紡錘體的方向,決定卵裂面的形成,決定卵裂類型。不僅如此,在一些特別適宜的卵子還可看到形成各種器官的物質在卵子中已經有了布局,卵裂之後各個裂球與將要形成的器官有一定的對應性。所有這些都提示,細胞核在遺傳潛能上是等同的,只是在以後的發育中,通過細胞質或細胞間的相互作用才受到不同的調節。
對於細胞核的作用也有了充分的估價。1887年德國實驗胚胎學家T.H.博韋里使海膽卵子被兩個精子受精,根據染色體在各個卵裂球中的分配以及各個卵裂球的發育情況,認為各個染色體有質的不同,染色體是有個性的。利用海膽卵子,T.H.摩爾根1896年完成了人工孤雌生殖──卵子不經受精也可發育。使不具細胞核的卵塊受精或用異種精子受精,研究細胞質及細胞核各自在發育中的作用,觀察到所產生的幼蟲都顯示父方的特徵。這些都說明細胞核的重要性。總括當時的成就,1883年德國胚胎學家W.魯曾經表達這樣的設想:“不僅染色體,而且每一染色體的各個部分,對於決定個體的發育、生理和形態可能都是重要的。”1887年德國動物學家A.魏斯曼提出種質的假說。雖然這個假說被後來的實驗研究推翻了,但是在假說中提出的決定子與後來的基因之間是有某些思想上的聯繫可尋的。
為解決胚胎學的問題,還為細胞學提供了重要的實驗方法,這就是組織培養。美國胚胎學家R.G.哈里森在1907年為了研究神經纖維的生長創立了體外培養的方法,後來被美國生理學家A.卡雷爾接過去,發展成專門的技術。30年代之後越來越顯示出它的重要性,到今天,不僅是研究活細胞的各方面,甚至對許多其他學科來講也是必不可缺的技術。
遺傳學的影響。1900年重新發現G.J.孟德爾的研究成就后,遺傳學研究有力地推動了細胞學的進展。美國遺傳學家和胚胎學家T.H.摩爾根研究果蠅的遺傳,發現偶爾出現的白眼個體總是雄性;結合已有的、關於性染色體的知識,解釋了白眼雄性的出現,開始從細胞解釋遺傳現象,遺傳因子可能位於染色體上。細胞學和遺傳學聯繫起來,從遺傳學得到定量的和生理的概念,從細胞學得到定性的、物質的和敘述的概念,逐步產生出細胞遺傳學。
1920年美國細胞學家W.S.薩頓進一步指出遺傳因子和染色體行為間的平行現象,必然意味著遺傳因子位於染色體上,並且提到,如果兩對因子位於同一染色體上,它們可能按照,也可能不按照孟德爾規律遺傳,預示了連鎖的概念,加深了關於成熟分裂尤其是關於染色體配對、染色體交換的研究。
此外,發現了輻射現象(X射線、鐳輻射、紫外線)、溫度能夠引起果蠅突變之後,因突變的頻率很高更有利於染色體的實驗研究。輻射之後引起的各種突變,包括基因的移位、倒位及缺失等都可在染色體中找到依據。利用突變型與野生型雜交,並且對其後代進行統計處理,可以推算出染色體的基因排列圖。
多線染色體的發現則打開了染色體研究的新途徑。在斷定了多線染色體就是加粗的,已配對的染色體之後,一方面對它的結構進行細緻的研究,發現了染色線上的染色粒,許多相鄰的染色粒聚集成帶區,染色線雖然不易看清楚,但是如果染色適宜或是在紫外光下可以看到它們不是筆直平行排列,而是很疏鬆的螺旋狀。另一方面可以把根據連鎖群推算出來的染色體上的基因排列圖利用所謂的唾腺方法和形態學的染色體圖吻合起來;雜交實驗和細胞的形態學觀察可以完善地互相印證,可以在多線染色體上更具體地確切地看到基因排列的情況,每個帶區實際上不只含有一個基因。不僅如此,有些突變是由於基因的位置效應例如棒眼突變型(bar-eye) 就是先在多線染色體上取得證據的。
在尋找遺傳的物質基礎的推動下,染色體的研究在面上鋪展開了,不僅用於遺傳研究的材料,許多其他動、植物物種(有人統計大約有12000種維管植物和500多種哺乳動物)的細胞分裂(減數分裂)、染色體行為、染色體圖譜都被研究過。同一屬中的物種,染色體的數目往往是一致的;但是同一科中的物種或者數目不等,或者這一屬的是另一屬的倍數(多倍性)。同一個體的各個染色體,粗看似乎無大差別,但是仔細檢查是有不同的,因此可以精確說出一個物種的染色體的數目、形狀以及各個染色體的大小,並且能夠把它們編號排隊。可以比較親緣關係較近的不同物種的染色體,由此尋找物種的進化關係;核型的研究指出相近的物種,其染色體數目可能完全一致,但是也可能出現十分明顯的差別,在後一種情況經過仔細研究總可找出原始形式,和由此派生出的各種形式。在植物已經知道有三種突變:多倍性、一個染色體斷裂成幾個小的或者相反的幾個小染色體集裝成一個大的以及某對染色體的倍增。這三種突變有時會和亞種及種的形成有關。此外,植物的多倍性的研究導致使用各種方法,例如化學物質、溫度、輻射等誘導多倍性的產生,在某些植物已經獲得應用的價值。
廣泛開展的性染色體形態的研究,也為雌雄性別的決定找到細胞學的基礎。有的動物是XX、XY型,有的是ZZ、ZW型。
細胞生理學的影響。在這個階段用實驗方法研究細胞其他部分的功能,沒有得到使人滿意的結果。用顯微鏡觀察不到細胞膜,只能根據細胞質與外界的物質交換判斷它的存在,以及某些物質的通透,藉以判斷它的某些功能。由於一般地說來脂溶性的物質易於進入細胞,曾經推測細胞膜可能由脂類或者脂類的小孔組成。也曾由於分子量不同的物質進入細胞的難易不同──分子量越大越難進入;推測細胞膜像是一個過濾層,它的小孔阻止大分子進入細胞。此外,曾根據電解質,例如陽性離子和陰性離子對細胞的通透,以及細胞環境的酸度可以影響、以致改變陽性和陰性離子的通透,提出電荷假說以解釋細胞的通透性這一極其複雜的過程。至於對於固體顆粒的吞噬作用,通過模擬實驗,例如變形蟲對氯仿滴的吞噬,認為這是由於細胞對異物的表面比對周圍環境有更大的粘著性,通過粘著引起細胞膜表面張力的局部變化以致異物被吞入。
上述這些設想,即使在那時看來,在通透性方面細胞膜都是被動的;但是細胞還能夠逆著擴散梯度或濃度梯度主動地攝入或排出某些物質。因此也曾設想,細胞膜中可能存在著需要能量的過程,它們對於這些過程有重大意義,但當時還沒有資料。
那時對細胞呼吸的理解主要局限於食物經過各種酶的作用產生出熱量。由於知道了在這過程中的幾種酶,例如某些脫氫酶、氧化酶、細胞色素a、c、b等,因而了解到食物在細胞中的燃燒不是通過一次突然的氧化把全部能量以熱的形式釋放出去,而是逐漸地通過一個個小的階段,一步步地獲得並且利用少量的能量。這種過程由於許多種酶作為轉移氧、接受氫、氧化還原體系等加入到總的呼吸過程中才能夠進行下去,並且得到微細的調節。
其他學科的影響。在20世紀40年代初期,其他學科的技術方法相繼被用於細胞學的研究,開闢了新的局面,形成了一些新的領域。首先是電子顯微鏡的應用產生了超顯微形態學。
比利時動物學家J.布拉謝從胚胎學的問題出發,利用專一的染色方法(Unna,Feulgen)研究核酸在發育中的意義。差不多與此同時,瑞典生化學家T.O.卡斯珀松根據各種物質對一定波長的吸收,創建了紫外線細胞分光光度計,來檢測蛋白質、DNA和RNA這些物質在細胞中的存在。如果說,前者根據染色可以做到定性,後者則根據吸收可以做到定量。實質上是他們的工作引起人們對核酸在細胞生長和分化中的作用的重視。在他們工作的基礎上發展起了細胞化學,研究細胞的化學組成,可以和形態學的研究相互補充,對細胞結構增加一些了解。
用多線染色體進行分析,在紫外光下拍照表明染色粒以及核仁含有DNA,相反地染色線只含很少,或者甚至沒有。用蛋白酶(可能不純)消化可以使它們溶解,因此曾誤認為染色線是蛋白質構成的。除此而外,還可根據紫外吸收光譜精確測定染色體段落(常染色質和異染色質)某些氨基酸的百分比。常染色質的段落似乎含有較多高分子量的球蛋白類型的蛋白質,而異染色質段落則含有較多低分子量的組蛋白類型的蛋白質。
20世紀40年代開始逐漸開展了從生化方面研究細胞各部分的功能的工作,產生了生化細胞學。首先使用了勻漿──在適合的溶液中把細胞機械地磨碎──和差速離心的辦法,除細胞核而外還可以得到線粒體、微粒體和透明質等幾部分。對它們分別地進行研究了解到一些物質和酶的存在和分佈以及某些代謝過程在什麼部位進行。分離得比較成功的是線粒體,因為用電子顯微鏡已經測量出它們的大小並且粗略地了解到在這種細胞器里進行的生化過程,認識到它們對能量代謝的重要性。微粒體曾經被誤認為是一種細胞器。後來知道,這是在當時的分離條件下的產物,是由核糖體和少量內質網組成的複合體。雖然如此,關於線粒體和微粒體這樣一些研究指出,許多基本的生化過程是在細胞質而不是在細胞核里進行的。這樣的方法結合著深入的形態學研究導致對細胞中的過程有越來越深刻的了解。
放射性同位素的應用為研究細胞中的代謝過程開闢了新的途徑。從它們的參入可以精確追蹤細胞內物質的合成、運輸、以及儲藏物的利用。例如用這方法顯示出磷的化合物不是在有絲分裂時,而是在間期、在分裂開始前不久參入,然後被分配到子細胞核。從這樣一些以及用其他同位素得到的結果,可以推斷細胞中的一些重要物質的運轉。
雖然在20世紀30年代組織培養就有了較大的發展,但是只能培養組織塊,還不能培養正常組織的單個細胞,而且還沒有充分顯示出它的重要性。利用培養的細胞可以研究許多在整體中(在原位)無法研究的問題,例如細胞的營養、運動、行為、細胞間的相互關係等。幾乎各種組織,包括某些無脊椎動物(墨魚、海鞘、果蠅等),都被培養過。在良好的培養條件下從組織塊長出的各種細胞,其生長情況不同。從形態上基本上可以分為三種類型,上皮、結締組織和遊走細胞(如淋巴細胞、單核細胞和巨噬細胞)。有時候培養細胞會顯示正常組織在有機體中表現不出的特徵,例如如果培養基中含有增強表面活性的物質,多種組織的細胞可以獲得吞噬的能力。但是它們仍保持特有的性質和潛能,因為如果改變培養環境或者移回到動物體內原來的部位便仍可照原樣生長。
值得一提的是在培養中的成纖維細胞的生長也受底質的影響。在一般情況下它們呈輻射狀、漫無目的地從組織塊長出。但是如果人工地使培養基處於一定方向的張力之下,或人工的在底質上制出痕迹,細胞就會沿張力的方向或沿著痕迹生長出去。這個現象也許可以用來解釋在整體中結締組織和肌腱的功能適應──它們總是在張力的方向生長、分化。
可以看出,對於細胞的研究,在使用電子顯微鏡后在亞顯微結構方面的深入,以及在應用生化技術后在功能方面的深入,已經在為細胞生物學──在分子水平上研究細胞的生命現象──的形成創造了條件。所以在後來,在分子遺傳學和分子生物學優異的成就的影響之下,細胞生物學這一新的學科很快地形成了。
在實驗室中,細胞有絲分裂試驗應注意以下技術環節:
一、紡錘體阻斷劑(Spindle inhibitor)
在有絲分裂過程中,隨著紡錘絲的形成,染色體被牽引到一起難以觀察其形態。紡錘體的形成在於細胞質和紡錘體成分的粘度之間的平衡,因此,改變細胞質的粘度,即可破壞紡錘體形成,從而使得染色體均勻散開,且染色體縊痕區更為清楚。
在培養中使用的紡錘體阻斷劑為秋水仙素,在終止培養前加入適量秋水仙素,使正在分裂的細胞停留在中期,以獲得大量分裂相供分析之用。秋水仙素的濃度範圍比較寬,一般使用濃度0.05—0.8微克/毫升,在終止培養前處理2—4小時。但在實際工作中需要藉助濃度和處理時間來控制染色體的收縮程度。秋水仙素作用時間越長,被阻斷的中期分裂相越多,但是染色體也越凝聚和收縮,所以還視各實驗室經驗而定。
二、低滲液(hypotonic solution)
低滲液是指滲透壓和離子強度均低於正常細胞生理條件的溶液,例如水、低滲的檸檬酸鈉或氯化鈉、甘油磷酸鉀(0.65M)、氯化鉀(0.075M)等。低滲效果取決於低滲液的化學組成、低滲的溫度和處理時間。低滲處理是憑藉反滲透作用使細胞膨脹染色體鋪展,同時可使粘附於染色體的核仁物質散開,以便能在一個平面上觀察所有染色體形態。
實驗室中一般選用0.075M KCl為低滲液(具體情況取決於實際操作,鑒於實驗的連續性和穩定性,本實驗室採用0.35%KCl),其優點有:①染色體輪廓清楚,可染色性強,染色時間短,②用於顯帶染色時能充分顯示帶型特點。
低滲處理為37℃,15-25分鐘,以預設實驗條件為準。
三、固定液
固定液的重要特性是能迅速穿透細胞,將其固定並維持染色體結構的完整性,還要能夠增強染色體的嗜鹼性,達到優良染色效果。
單純的固定液一般難以達到這些要求,因此在實驗中使用兩種混和的固定液。由於Carnoy首先使用的甲醇和冰乙酸混合液而稱的卡諾氏固定液是效果良好的固定液。Carnoy固定液(甲醇:冰乙酸=3:l)每次使用前需臨時配製,長時間放置影響固定效果,固定時間15分鐘至24小時,冰箱、室溫均可。必要時可改變甲醇和冰乙酸的比例,冰乙酸比例增加,利於細胞膨脹、染色體鋪展,但易導致細胞破裂、染色體散失。
四、滴片
滴片使細胞懸液從一定高處落在載玻片上,淋巴母細胞膜破裂,染色體分散開。載玻片可用冰片和乾片,效果均好。滴片后需空氣乾燥。
五、Giemsa染色
Giemsa染料不是一種單一染料,而是天青、伊紅、甘油和甲醇的混合物,配製后原液儲存。在常規染色中,並不比其他染料優越,但在顯帶技術中,卻是無可比擬的。
Giemsa工作液在使用前臨時配製,濃度可在2.5-10%之間(原液2份加pH7.4磷酸緩衝液10-40份)。染色后,染色質呈紅色,細胞核是藍色。
基本信息
- 中文名
- 細胞學
- 外文名
- Cytology
- 適用領域
- 化學領域
- 學科分類
- 生物學