神經突觸
神經突觸
神經突觸,1896年C.S.Sherrington把神經元與神經元之間的機能接點命名為突觸,有兩類突觸:電突觸與化學突觸。
1896年C.S.Sherrington把神經元與神經元之間的機能接點命名為突觸(synapse),當時他雖然還不了解接點的形態學,但是他指出神經元與神經元之間是不連續的,而且推論有些突觸是興奮性的有些突觸是抑制性的。在20世紀30、40年代對於突觸之間是電學傳遞還是化學傳遞曾經發生過爭論。現在知道有兩類突觸:電突觸與化學突觸。
神經元之間化學傳遞的基本概念起源於哺乳動物內臟神經系統的研究。本世紀初,J.N.Langley和他的學生髮現腎上腺素的效應與刺激交感神經系統的效應十分相似。他的學生,T.R.Elliott甚至指出,腎上腺素可能是外周神經釋放的化學刺激物。後來H.H.Dale發現膽鹼及其衍生物對心臟、膀胱和唾液腺的效應與刺激副交感神經相似,特別是乙醯膽鹼最有效。Dale提出乙醯膽鹼、腎上腺素的作用與刺激兩類內臟神經的效應相似性的問題。Otto Loewi在1921年所做的實驗證明,刺激迷走神經釋放活性化學物質,抑制心搏。繼而證明,這種化學物質就是乙醯膽鹼。1936年Dale等人在刺激支配肌肉的運動神經后得到了神經釋放的乙醯膽鹼,因而把化學傳遞的假說推廣到全部外周神經系統。證明乙醯膽鹼是神經肌肉接點的神經遞質后,直到1952年中樞神經系統的化學遞質說才被廣泛接受,而在7年之後,E.Furshpan和D.Potter又第一次清楚地證明了電突觸的存在。
Furshpan和Potter在1959年首先指出在螯蝦的可興奮細胞之間有電學傳遞。電學傳遞可以發生在中樞神經系統的細胞之間、平滑肌細胞之間、心肌細胞之間、感受器細胞和感覺軸突之間。一個電突觸的突觸前膜和突觸后膜緊緊貼在一起形成縫隙連接,電流經過縫隙連接從一個細胞很容易流到另一個細胞。向見圖的A細胞送入閾下電脈衝,引起這個細胞膜電位的變化。如果送入細胞A的電流相當一部分經過縫隙連接流入細胞B,就會相應地在B細胞引起可以察覺的膜電位變化。由於只有一部分流入細胞A的電流進入細胞B,所以細胞B的膜電位變化比細胞A的小。這種電突觸也同樣允許動作電位的局部電流通過,因此動作電位在縫隙連接之間傳播與在軸突上的傳播沒有什麼本質不同。因為兩者都依靠先行於動作電位的局部電流來興奮膜上的新區域。從一個細胞到另一個細胞的縫隙連接的電阻一般是對稱的,沒有方向性,但也有一些例外。例如螯蝦的大運動纖維之間的電突觸的電阻不是對稱的,有類似整流器的作用,電流向某一方向流動比向相反方向的容易。因此,一條軸突上的衝動可以引起第二條軸突的動作電位,但是第二條軸突上的衝動不能引起第一條軸突上衝動的發放。在電突觸的突觸前細胞和突觸后細胞的電位變化之間基本上沒有突觸停滯。這種突觸傳遞對缺氧、離子或化學環境的變化不敏感,這些因素不能阻斷動作電位。因此,電突觸與化學突觸相比較,由於它包含的步驟較少,在傳遞過程中實行控制、改造的機會也比較少。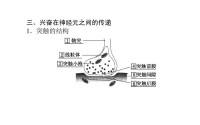
神經突觸
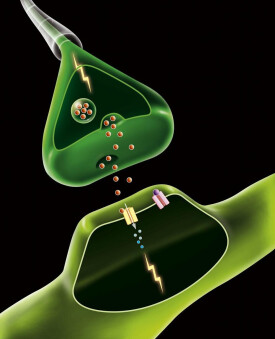
在許多動物(包括腔腸動物、環節動物、節肢動物、軟體動物、低等和高等脊椎動物)身上都發現在某些神經元之間存在縫隙連接和電學傳遞,但大多數突觸傳遞是化學傳遞,通過突觸前神經元的末梢分泌傳遞物質,使突觸后膜的離子通透性發生變化,產生突觸后電位。一般地說,化學傳遞比電傳遞有更大的可塑性,而且可以把比較小的突觸前電流放大成比較大的突觸后電流。
化學傳遞發生在突觸間隙之間。突觸前細胞膜與突觸后細胞膜之間的間隙寬約20納米,間隙中填充著粘多糖,把突觸前膜與突觸后膜“膠合”在一起。突觸前末梢包含幾百、上千個突觸小泡,直徑約40納米。每個小泡中含有104到5×104個遞質分子。當神經衝動傳到突觸前末梢,突觸前膜去極化,使鈣離子由膜外進入膜內,促使一定數量的突觸小泡與突觸前膜接觸,觸點融合併出現裂口,小泡內的化學遞質進入突觸間隙。遞質由於擴散而到達突觸后膜。遞質達到突觸后膜即與膜上的特殊的受體結合,改變突觸后膜對某些離子的通透性,使膜電位發生變化。這種電位變化叫做突觸后電位(postsynaptic potential)。
神經突觸
神經遞質,特別是中樞遞質的化學特性的研究還存在不少的困難,許多突觸的神經遞質還不能確定。要確定某一物質是某組織的遞質必須符合一些標準,如下列幾條:
(1)施用於突觸后膜時,它引起突觸后細胞的生理效應與突觸前刺激所引發的生理效應相同。
(2)突觸前神經元活動時必定釋放這種物質。
(3)它的作用必定被能夠阻斷正常傳遞的阻斷劑所阻斷。
表1列舉一部分神經遞質和它們的作用部位。
乙醯膽鹼是目前最熟悉的神經遞質。脊椎動物運動軸突末梢、脊椎動物植物性神經系統的節前末梢、副交感神經的節后末梢以及脊椎動物中樞神經系統某些神經元的突觸前末梢都釋放乙醯膽鹼。某些無脊椎動物的神經元的遞質也是乙醯膽鹼。乙醯膽鹼從軸突釋出與受體結合后就被突觸后膜上乙醯膽鹼酯酶水解成膽鹼和乙酸,終止了乙醯膽鹼對突觸后膜的作用。膽鹼被突觸前末梢吸收用以重新合成乙醯膽鹼(見圖)。
(乙醯膽鹼受體還可區分為兩類:一類是毒蕈鹼型受體(muscarinic receptor)(簡稱M型受體);一類是煙鹼型受體(nicotinic receptor)(簡稱N型受體)。M型受體存在於所有的副交感神經節后纖維支配的效應器上,還存在於交感神經節后纖維支配的汗腺,以及交感舒血管纖維支配的骨胳肌血管上。乙醯膽鹼與M型受體結合可以產生一種類似毒蕈鹼與之結合所產生的作用,包括心搏抑制、支氣管胃腸平滑肌和瞳孔括約肌收縮、消化腺分泌、汗腺分泌、骨骼肌血管舒張等。阿托品可以與M型受體結合阻斷乙醯膽鹼的作用。N型受體存在於神經肌肉接點的突觸后膜與內臟神經節(交感、副交感神經節)的突觸后膜上。乙醯膽鹼與N型受體結合可以產生一種類似小量煙鹼與之結合所產生的作用,也就是骨骼肌和節后神經元的興奮。箭毒可以與神經肌肉接點突觸后膜上的N型受體結合阻斷乙醯膽鹼的作用;六烴雙胺可以與交感、副交感神經節中突觸后膜上的N型受體結合阻斷乙醯膽鹼的作用。
去甲腎上腺素、多巴胺和5-羥色胺是結構相近的化合物。某些脊椎動物和無脊椎動物的神經元存在這類化合物。它們大多數集中在神經末梢。去甲腎上腺素是交感神經系統節后細胞的興奮性遞質。上圖表示去甲腎上腺素和腎上腺素的生物合成途徑。
上圖表示在腎上腺素突觸中的化學變化。去甲腎上腺素由突觸前末梢釋出后,與突觸后膜上的腎上腺素受體結合發揮生理效應,大部分被末梢重新吸收加以利用,小部分被單胺氧化酶降解,還有一部分被甲基移位酶失活。終止乙醯膽鹼的作用靠膽鹼酯酶水解乙醯膽鹼,而終止去甲腎上腺素的作用主要靠末梢對遞質的重新吸收。腎上腺素受體也可分為α型腎上腺素受體(簡稱α受體)與β型腎上腺素受體(簡稱β受體)。去甲腎上腺素、腎上腺素與α受體結合引起效應器的興奮,但也有抑制的情況,如小腸平滑肌;與β受體結合則引起效應器的抑制,但對心臟的作用是興奮,部分腎上腺素受體的分佈與效應見表。
某些氨基酸,如谷氨酸,是脊椎動物中樞神經系統興奮性突觸和昆蟲、甲殼動物興奮性神經肌肉接點釋放的遞質。γ-氨基丁酸(Gamma aminobutyric acid,GABA),NH2—CH2—CH2—CH2—COOH,是脊椎動物中樞神經系統(大腦皮層、小腦)的抑制性遞質,起著很重要的作用。甲殼動物的抑制性運動突觸的遞質也是γ-氨基丁酸。
神經肽(neuropeptide)是上述少數“經典”遞質以外的一組多肽分子,它們在神經系統中產生和釋放,作為遞質或影響突觸傳遞的突觸調質(synanpic modulator)發揮作用。第一個神經肽是U.S.von Euler和J.H.Gaddum在1931年發現的。當他們檢定兔腦和小腸提取液中的乙醯膽鹼時發現,這種提取液引起遊離腸段的收縮類似乙醯膽鹼的作用,但是這種收縮不能被乙醯膽鹼拮抗劑所阻斷。他們發現收縮是由一種多肽引起的,命名為P物質(substance P)。此後一系列的神經肽相繼在脊椎動物的中樞神經系統、外周神經系統、植物性神經系統以及無脊椎動物的神經系統中被發現。有趣的是有些神經肽最初是在內臟中發現的,如胃腸激素中的高血糖素、膽囊收縮素。近年來用熒光抗體免疫標記法可在組織切片中定位神經肽。現在已知的神經肽有幾十種,包括P物質、腦啡肽(enkephalin)、血管活性腸肽(vasoactive intestinal polypeptide,VIP)、抗利尿素、催產素、促腎上腺皮質激素釋放因子等等。有些神經肽既可作為神經遞質又可作為神經激素發揮作用,如同去甲腎上腺素一樣。近年來還發現有些神經肽可以同經典遞質如乙醯膽鹼、去甲腎上腺素共存於神經末梢中,作為經典遞質的協同遞質(cotransmitter)釋放出來。
腦啡肽和內啡肽(endorphin)是引人注意的兩類神經肽,因為它們有鎮痛和產生阿片樣欣快感的作用。與其它的神經肽的發現有所不同,在體內首先發現的是阿片受體(opioid receptor)。在1973年有三個實驗室獨立發現在哺乳動物腦中存在阿片受體,這些受體能與阿片樣物質結合併啟動其效應。阿片受體的存在預示著體內也有內源性阿片樣物質,因為阿片受體的天然作用不會是與罌粟等類植物的提取物相互作用,而是與體內某些阿片樣物質相互作用。幾年後在哺乳動物腦中發現具有阿片樣活性的肽類,阿片肽(opioid peptide)。阿片肽的大小相差頗大,從5個氨基酸的腦啡肽到31個氨基酸的β-內啡肽,但它們都有5個共同的氨基酸序列,即酪氨酸-甘氨酸-甘氨酸-苯丙氨酸-甲硫氨酸(或亮硫氨酸)。這一序列是阿片肽的標誌,也是它們與阿片受體結合併表現出阿片樣藥理活性所必需的。
Dale於1935年提出,神經元是一個統一的整體,它的各個末梢所釋放的遞質應是同樣的。1957年J.Eccles進一步概括為一種神經元只釋放一種遞質,被稱為Dale原則。因之神經元就以所釋放的遞質命名,如乙醯膽鹼神經元、腎上腺素神經元等。近年來的研究表明,一種神經元可以包含不止一種遞質,如在經典遞質之外還有神經肽。在支配貓唾液腺的頜下神經節的部分細胞中有血管活性腸肽(VIP)與乙醯膽鹼共存。刺激支配頜下腺的副交感神經可從頜下腺的靜脈血中檢出乙醯膽鹼和血管活性腸肽,說明神經末梢釋放兩種遞質。低頻電刺激(2赫)引起血管舒張和唾液分泌,這些作用可以用毒扁豆鹼增強和阿托平阻斷,表明是乙醯膽鹼的效應。但高頻電刺激(15赫)引起的血管舒張不被阿托平阻斷,這是血管活性肽的效應。
近年來的研究表明,除神經遞質在神經調節中起作用外,還有神經調質(neuromodulator)也在發揮作用。神經調質是神經細胞和某些內分泌細胞所釋放的一些物質,它們不直接引起所支配細胞的機能變化,而是調製突觸前末梢釋放經典遞質的活動以及突觸后細胞對遞質的反應。