Toll樣受體
1980年發現的蛋白質分子
新近研究發現,TLR能結合機體自身產生的一些內源性分子(即內源性配體)。免疫佐劑可增強抗腫瘤免疫,其分子和細胞機製得到進一步闡明TLR也在其中扮演重要角色。由於腫瘤在發生髮展過程中可以產生一些能被TLR識別的內源性配體,所以TLR在腫瘤免疫監視中可能發揮了一定作用。
Toll樣受體(Toll-like receptors,TLR)是參與非特異性免疫(天然免疫)的一類重要蛋白質分子,也是連接非特異性免疫和特異性免疫的橋樑。TLR是單個的跨膜非催化性蛋白質,可以識別來源於微生物的具有保守結構的分子。當微生物突破機體的物理屏障,如皮膚、粘膜等時,TLR可以識別它們並激活機體產生免疫細胞應答。
早在19世紀人們了解到微生物致病的概念后,就想到多細胞生物體中應該存在這樣的分子,它們能夠識別微生物特有的分子,從而識別入侵的微生物。早在100多年前,人們就開始尋找這樣的分子。德國著名細菌學家科赫的弟子理查德菲佛(Richard Pfeiffe)創造了”內毒素“一詞來稱呼革蘭氏陰性細菌中能夠造成動物發熱和休克的一種成分。後來人們發現這種物質是大多數革蘭氏陰性細菌產生的脂多糖(LPS)。人們又發現,另外一些分子,如細菌脂肽、鞭毛蛋白、非甲基化DNA等,能夠激發宿主的保護性應答,但是如果這樣的應答持續過久或者強度過大,就會造成傷害。於是人們從邏輯上推定機體中必定存在這樣的分子的受體,它們可以向機體發出存在感染的警報。然而時隔多年人們卻沒有找到這樣的受體。
1980年,Nusslein-Volhard等在研究果蠅胚胎髮育過程中發現有一個基因決定著果蠅的背腹側分化,將其命名為Toll基因。1988年,Hashimoto等人發現Toll基因編碼一種跨膜蛋白質,並闡明了Toll蛋白的結構。1991年,Gay等人發現,Toll蛋白在結構上與哺乳動物中一種天然免疫功能分子——白細胞介素1受體(IL-1R)具有同源性:二者的細胞質部分相似。這第一次提示了人們Toll可能和免疫有關。1994年,Nomura等人首先報道了人中的Toll樣受體。然而當時Toll的免疫學功能沒有得到闡明,所以人們仍然認為Toll樣受體是和哺乳動物的發育有關的。不過,兩年之後的1996年,Jules A.Hoffmann和他的同事們發現Toll在果蠅對真菌感染的免疫中起著重要作用,從而確立了Toll的免疫學意義。翌年,Charles Janeway和Ruslan Medzhitov闡明了一種Toll樣受體(後來被命名為TLR4)能夠激活與適應性免疫有關的基因。Bruce A.Beutle隨後發現TLR4能夠探測LPS的存在。後來他們又發現,如果使小鼠中的TLR4突變而喪失功能,小鼠不會對LPS起反應。
後來,科學家們用基因打靶的方式使其他各種TLR喪失功能進行了研究。結果人們認為,每種TLR可識別不同的一類分子。
自此,人們不但揭示了Toll樣受體在免疫學上的意義,也揭開了100多年前人們尋找的那個謎底。
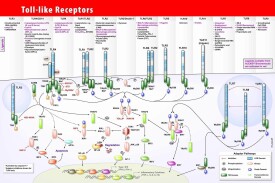
所有Toll樣受體同源分子都是Ⅰ型跨膜蛋白,可分為胞膜外區,胞漿區和跨膜區三部分。
Toll樣受體胞膜外區主要行使識別受體及與其他輔助受體(co-receptor)結合形成受體複合物的功能。Toll樣受體的胞漿區與IL-1R家族成員胞漿區高度同源(IL-1R介導的信號傳導系統和機制與果蠅類似),該區稱為Toll-IL-1受體結構域(Toll-IL-1 receptor domain,TIR結構域)。TIR具有嗜同性相互作用(homophilic interaction),藉此來募集下游含有TIR的信號分子,組成信號複合體。但是二者胞外部分不相關,TLR胞膜外區為有17個~31個亮氨酸富集的重複序列(Leucine rich repeats,LRRs),並且都含有3個胞外段輔助蛋白即MD-1、MD-2和RP105,參與對疾病相關分子模式(PAMP)的識別;而IL-1R為Ig樣的結構,在哺乳動物中,一些胞漿蛋白也存在TIR域,如2種信號接受蛋白,MyD88和TIR域的接受子蛋白(TIR domain-containing adaptor protein,TIRAP),二者在TIR信號傳導中起作用。
目前,在哺乳動物及人類中已經發現的人TLRs家族成員有11個。其中了解比較清楚的有TLR2,TLR4,TLR5和TLR9。人的TLRs家族基因定位分別是定(TLR1,2,3,6,10)4號染色體,9號染色體(TLR4),1號染色體(TLR5),3號染色體(TLR9),x號染色體(TLR7,8)。
根據TLRs細胞分佈特徵,可將其分為普遍存在型(TLR1)、限制存在型(TLR2、TLR4、TLR5)及特異存在型(TLR3)3類。
根據染色體的位置、基因結構和氨基酸序列,人的TLRs受體可以分為5個亞科,即TLR2,TLR3,TLR4,TLR5和TLR9。TLR2亞科有TLR1,TLR2,TLR6和TLR10;TLR9亞科有TLR7,TLR8和TLR9;TLR3,TLR4和TLR5各自形成一個亞科。
TLRs分佈的細胞多達20餘種,Muzio M 等對TLR1-TLR5表達於人類白細胞的研究中發現,TLR1能在包括單核細胞,多形核細胞,T、B淋巴細胞及NK細胞等多種細胞中表達,TLR2、TLR4、TLR5隻在髓源性細胞(如單核巨噬細胞)上表達,而TLR3隻特異性表達於樹突狀細胞(dendritic cells,DC)。
Toll樣受體在天然免疫中的識別作用
●TLR4可以識別革蘭氏陰性菌脂多糖(LPS),還可識別宿主壞死細胞釋放的熱休克蛋白(heat-shockproteins,HSP),體內類肝素硫酸鹽和透明質酸鹽降解的多糖部分以及局部的內源性酶的級聯活化反應也可激活TLR4。
●TLR2的配體較TLR4的廣泛,包括脂蛋白,脂多肽,脂肪酸(LTA)阿拉伯甘聚糖(LAM)及酵母多糖等。
●TLR3特異識別病毒複製的中間產物ds-RNA,從而激活NF-кB和干擾素IFN-β前體。DoyleS E等證實,抗TLR3單克隆抗體能抑製成纖維細胞IFN-β的產生。Christopher A等證實TLR3還具有調控鼻病毒對人支氣管細胞感染的能力,這也說明了TLR3在宿主抵抗活病毒中發揮重要的作用。
●TLR7識別咪喹啉家族低分子量的咪唑莫特、R-848和R-847等。
TLR7、TLR8和TLR9高度同源,與其他TLR不同,它們在細胞內涵體中起作用,吞噬和包膜溶解后結合它們的配體,可識別微生物的核酸。
另外,TLR對配體的識別,不同類型的TLRs可以組合,從而識別不同的PAMPs,如TLR1與TLR6可以協同TLR2對不同的PMAPs分子進行組合識別;TLR7可能同TLR9組合來介導CpG激活免疫細胞。其中TLR4/TLR4和TLR9/TLR9是以同源二聚體的形式進行;而TLR2/TLR4、TLR2/TLR6和TLR7/TLR8為異源二聚體,還有的二聚體中有一個亞單位尚未確定,如TLR3/TLR、TLR5/TLR。
Toll樣受體在獲得性免疫系統中的作用
首先,Toll樣受體在獲得性免疫中的具有識別作用。機體最強的抗原呈遞細胞——樹突細胞可表達TLR。藉助TLR識別LPS、GpG-DNA、肽聚糖、脂蛋白以及分支杆菌的細胞壁成分等具有PAMP的分子,樹突細胞被活化而成熟,提供獲得性免疫的共刺激信號。因此TLR是微生物成分引起樹突細胞活化的橋樑。
具體地說,TLR3、TLR7/TLR8、TLR9在病毒核酸成份的刺激下,誘導機體產生I型干擾素,後者發揮抗病毒免疫作用。TLR2和TLR4激活DC后產生不同的細胞因子和化學激活因子。TLR4主要產生IL-12 p70,IFN-γ介導蛋白(IP-10)及轉錄IFN-β。TLR2刺激則優先表達IL-8和IL-23。這些可溶性細胞因子誘導T輔助細胞向有利於殺滅病原的方向分化產生細胞免疫應答或體液免疫應答。尤其是IL-12和IP-10能夠刺激T細胞產生IFN,促使Th細胞分化為Th1細胞。如果缺乏IL-12則分化為Th2細胞。
另一方面,TLR2優先激活p19的轉錄。p19是近期才被證明的一種可以和p40形成雜二聚體的蛋白,與IL-12具有相似的作用。TLR4激動劑可刺激大量產生IP-10。IP-10是不同細胞對IFN-γ、微生物成分應答后產生的CXC化學激活因子,可對單核細胞、NK細胞產生化學吸附作用,更重要的是Th1型細胞優先表達IP-10受體CXCR3,但TLR2刺激DC不能表達IP-10。