軟骨藻酸
軟骨藻酸
軟骨藻酸(Domoic acid),非蛋白氨基酸,是由長鏈羽狀硅藻代謝產生的一種強烈的神經毒性物質,能導致短期記憶功能的長久性損害。自從1987年加拿大中毒事件后開始被認識。毒性作用機制可能為:通過谷氨酸與內源性神經遞質協同,胞內鈣離子超載使信號轉導紊亂;代謝紊亂使神經元不具有足夠的能量維持正常的靜息膜電化學梯度。
軟骨藻酸( ,)氨基酸,污染貝類離、提取獲擬菱形藻+尿素產量軟骨藻酸。興奮脯氨酸衍毒素,浮游植物代謝的產物,可以在被藻類污染的海洋食物特別是貝類中檢測到,其結構與紅藻氨酸和谷氨酸相似,是紅藻氨酸受體的興奮劑。主要由某些擬菱形藻屬和菱形藻屬的海洋硅藻產生,人們食用毒化的貝類,可引起記憶喪失、眩暈、昏迷甚至死亡等癥狀,根據這種毒素的中毒特徵,被命名為記憶喪失性貝毒(Amnesic Shellfish Poisoning,ASP)。
拿華王島岸食紫貽貝造毒亡件,造毒,亡。毒腹痛、腹瀉、嘔吐,伴嚴痛、憶喪、識混亂、昏迷癥狀。病癒憶喪達久。拿洋研究專鑒,毒質列擬菱形藻( )產軟骨藻酸( )。,拿、岸擬菱形藻產軟骨藻酸引危害件斷。
拿岸,貽貝中也檢測出了由偽優美擬菱形藻產生的DA。1991-1994年,太平洋海岸連續爆發由澳洲擬菱形藻形成的赤潮,由此造成魚貝類被DA嚴重污染。1991年,美國的太平洋海岸爆發了有毒擬菱形藻赤潮,加利福尼亞的蒙特利灣有100多隻褐色鵜鶘和鸕鶿吃了富集有DA的鳳尾魚而死亡,產毒藻為澳洲擬菱形藻。同年10-11月份,從華盛頓州及俄勒岡州產的竹鰠、加州產的殼菜及蟹類中也檢出高濃度的DA。1992年12月,日本從美國進口的蟹(Cancermagister)內臟DA陽性。1996年,墨西哥Baja地區,成百上千隻海鳥因食用體內富集高濃度DA的螃蟹、鳳尾魚、沙丁魚中毒死亡。1998年美國加利福尼亞州中部沿海發生400多頭海獅中毒死亡事件,存活的海獅中有神經功能失調癥狀,從海獅的體液中檢測出了高濃度DA。
由硅藻門(Baciilariophyta)、羽紋硅藻綱(Pennatae)、管殼縫目(Aulonoraphidinals)、菱形藻科(Nitzschiaceae)中的擬菱形藻屬(Pseudo-nitzschia)和菱形藻屬(Nitzschia)中硅藻的某些種產生。水中的蛤蜊、蚌類及魚均可富集水中藻類產生的軟骨藻酸,並對軟骨藻酸有較強的耐受力,軟骨藻酸經食物鏈富集后,將對所在地區的生態環境造成影響,並引起動物發病和死亡Walz等檢測了水中Pseudonitzschia australis藻含量及軟骨藻酸的濃度,當該藻類在水中的含量僅為4000個/L時,即可檢出軟骨藻酸。因此應在赤潮和水華髮生的前後,加強藻類及軟骨藻酸污染的檢測,並開展軟骨藻酸對中樞神經系統損害機理的研究,對預防和治療軟骨藻酸中毒是十分必要的。
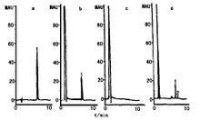
軟骨藻酸色譜
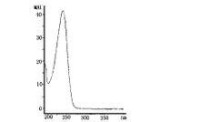
軟骨藻酸紫外光譜
測定貝類中軟骨藻酸的方法主要有高效液相色譜法、毛細管電泳法、小白鼠生物學試驗法等,其中高效液相色譜法是檢測貝類中DA公認的有效方法。高效液相色譜對貝毒素的分析應用最成功的是對軟骨藻酸的分析。一般條件下HPLC-UV可使軟骨藻酸得以分離並有效地檢測。為保證分析的回收率,在樣品處理時,應儘可能減少酸性溶液在空氣中的暴露時間。

軟骨藻酸離子掃描圖
軟骨藻酸的分子式為C15H21NO6,分子量為311.33。結構與紅藻氨酸和谷氨酸相似,是紅藻氨酸受體的興奮劑。純品為白色固體粉末,溶於水,微溶於甲醇,熔點223-224℃,在紫外光譜區最大吸收波長為242nm,在體積比為1:9的乙腈/水溶液和-12℃黑暗條件下可保持穩定一年左右。在常溫或光照下在鹼性溶液中不會降解,但它在酸性溶液(pH=3)中一星期降解50%。分子中含有三個羧基和一個仲氨基,羧基結構的pKa分別為2.10、3.72、4.97,氨基結構的pKa為9.82,因此它在溶液中的存在受pH的影響。軟骨藻酸共有A-H八種異構體。其中異構體A、B、C的毒性遠低於其餘五種。

不同貝類中軟骨藻酸含量的變化

軟骨藻酸標準質譜圖

軟骨藻酸在大馬哈魚體內分佈情況
雖然在1987年就報道了人類的ASP事件與DA有關,但到1991年九月才第一次明確發現有水產動物死亡與DA毒素有關。在貝類和魚類體內存在DA使得毒素有可能通過食物鏈進行傳播,即使DA沒有累積到致死劑量,也會對健康造成慢性影響。然而,尚無有效方法預測到DA有何慢性影響。對於DA的研究還有待於檢測技術的進一步發展。

軟骨藻酸二級質譜圖

軟骨藻酸作用細胞2h后氨基酸釋放色譜

軟骨藻酸作用細胞2h后遊離鈣濃度
人類的DA中毒反應主要根據1987年加拿大愛德華王子島DA中毒事件,患者在攝入DA數小時后出現噁心、嘔吐、腹絞痛、腹瀉、出血性胃炎和食慾減退等中毒癥狀和體征,嚴重者在出現胃腸道癥狀后數小時至3天內出現嚴重的頭痛、共濟失調、頭暈眼花、視覺障礙和記憶喪失、意識混亂、方向感喪失、失語、自主神經系統功能紊亂、不自主咀嚼、苦笑面容、肌陣攣、驚厥和昏迷等,並伴有永久性後遺症如記憶喪失和外周多發性神經病(peripheral polyneuropathy)。記憶缺失多發生在男性和年長者。四例死亡患者的神經病理學研究結果顯示,大腦的海馬和杏仁核的神經元損傷嚴重。用從導致中毒的貝類中提取的DA進行體外實驗,結果發現對培養的人神經元的毒性強於DA純品。DA作為驅蛔蟲藥的服用劑量是20mg/人。1987年導致加拿大人類中毒爆發事件中被測貝類中的DA含量為31-128mg/g,推算有癥狀病人DA的攝入量是60-290mg/人。對貝體內DA的食用安全性研究發現,貝組織中DA量達到40mg/kg時可引起食用者中毒,150mg/kg時有致死危險。人類通過膳食可耐受的最大限量為20mg/kg。

擬菱形藻的光學電子圖譜
擬菱形藻光學電子圖譜參考文獻。
擬菱形藻屬已有12種被報道可產生軟骨藻酸,其中澳洲擬菱形藻(P.australis)、成列擬菱形藻(P.seriata)與多列擬菱形藻(P.multiseries)的產毒性已得到廣泛的認可。而其他9種:靚紋擬菱形藻(P.calliantha)、細弱擬菱形藻(P.cuspidata)、擬柔弱擬菱形藻(P.pseudodelicatissima)、多紋擬菱形藻(P.multistriata)、尖刺擬菱形藻(P.pungens)、柔弱擬菱形藻(P.delicatesima)、P.fraudulenta、P.turgidula和P.galaxiae的毒性在不同地域則表現不同,擬菱形藻也同樣廣泛分佈於中國沿海,自南到北已報道的擬菱形藻有澳洲擬菱形藻、細弱擬菱形藻、柔弱擬菱形藻、多列擬菱形藻、多紋擬菱形藻、尖刺擬菱形藻、擬柔弱擬菱形藻、成列擬菱形藻、中華擬菱形藻、P. subpacifica,P. subfraudulenta,P. turgidula共12種。其中有9種是潛在產毒種,在非中國海區均有產毒報道,它們是:澳洲擬菱形藻、細弱擬菱形藻、柔弱擬菱形藻、多列擬菱形藻、多紋擬菱形藻、尖刺擬菱形藻、擬柔弱擬菱形藻、成列擬菱形藻、P. turgidula。雖然陳西平等(2001)檢測到中國渤海等海域部分海產品中存在低濃度的軟骨藻酸,李大志等(2002)也報道大連海域黑石礁省區的扇貝受到了軟骨藻酸的污染,但是尚無關於中國海域擬菱形藻產生軟骨藻酸的直接相關報道。
在以下區域的貝類樣品中發現有軟骨藻酸的存在:北美、紐西蘭、日本、丹麥、蘇格蘭、法國、西班牙、葡萄牙。雖然有毒擬菱形藻在世界各地的海岸都有分佈,但在不同地區是有不同種的擬菱形藻(或混合種)佔優勢。(加拿大的愛德華王子島東部是由P.multiseries 佔優勢;加拿大的Fundy灣是P.pseudodelicatissima;而在北美的西海岸則是P.multiseries,P.australis,和P.pseudodelicatissima共存;在荷蘭是P.multiseries和P.delicatissima;丹麥是P.seriata;西班牙是P.australis;紐西蘭是P.australis和P.delicatissima。)而且這些種在有時在某地有毒性,在另一地則會無毒。(如P.pseudodelicatissima在加拿大的Fundy灣;P.delicatissima 源自加拿大東部,現在在丹麥和紐西蘭也發現有毒;P.seriata在丹麥。)雖然極為稀少,但仍然存在著有毒的P.pungens 菌株(只在太平洋中出現)以及無毒的P.multiseries 菌株,必須用最靈敏的檢測技術才能確定是否有毒性。已知P.multiseries赤潮通常發生在較寒冷的季節(秋季到春季),而P.pungens,P.pseudodelicatissima和P.australis赤潮多發生在溫暖的季節。Rhodes等在1996年發現紐西蘭某地生長的尖刺擬菱形藻(P.pungens)能產生DA,而該藻在紐西蘭的其它地區卻不產毒;1994年Lundholm等發現原被認為不產毒的成列擬菱形藻在歐洲海域卻產毒;而在加拿大Fundy灣等地發現產毒的偽優美擬菱形藻在澳大利亞的Tasmanian和Victorian海域卻是無毒藻。共發現擬菱形藻屬20多個種,可產毒者有7種,包括多列擬菱形藻、偽優美擬菱形藻、澳洲擬菱形藻、優美擬菱形藻(P.delicatissima)、成列擬菱形藻、尖刺擬菱形藻(P.pungens)和虛假擬菱形藻(P.fraudulenta)。其中多列擬菱形藻、偽優美擬菱形藻和澳洲擬菱形藻為普遍認同的3個產毒藻種。
有報道東南亞諸島紅藻(C.armata)中DA的含量隨季節變化而異,最高可達1000mg/kg,從該海域採集的其它紅藻(如Jania capillacea、Coelothrix irregularis)中也含有大量的DA。日本產的貝類中至今尚未檢出DA,可能是日本的貝類被DA污染的途徑與北美和加拿大不同所致。公認的魚貝類被DA污染的原因是生物鏈外因學說,即與赤潮發生有關。水中的貝類和魚類對DA有較強的耐受力,它們可以富集藻類產生的DA,再經食物鏈的傳遞對所在地區的生態環境造成影響,人類食用被DA污染的海產品后即可引起中毒。在中國海域共發現擬菱形藻屬的5個種,分別是:中國擬菱形藻(P.sinica)、亞太平洋擬菱形藻(P.subpacific)、多列擬菱形藻、偽優美擬菱形藻和尖刺擬菱形藻。其中偽優美擬菱形藻在廈門海域發現並大量存在,但是否產毒尚不清楚。中國對擬菱形藻屬的研究主要集中在尖刺擬菱形藻上,中國近海尖刺擬菱形藻分為尖刺型(f.pungens)和多列型(f.multiseries)兩個型,前者不產毒,後者產毒。尖刺擬菱形藻是中國沿海普遍存在的種類,並且是引發赤潮的重要種類,在大連、青島、黃海長江口、廈門港及南海各港灣都引起過赤潮。1990年6月以來,中國南海大鵬灣多次爆發過尖刺擬菱形藻赤潮。赤潮生態研究結果顯示,硅藻中的尖刺擬菱形藻和偽優美擬菱形藻是優勢藻種,而且由尖刺擬菱形藻引發的赤潮最頻繁,幾乎全年都有發生,多出現於夏季。1997年7月-1998年6月對大亞灣海域擬菱形藻種群的研究發現,大亞灣水域的優勢藻種為擬菱形藻屬的尖刺擬菱形藻和偽優美擬菱形藻,一年四季均有出現,其中秋季、春季為兩個高發期,最高頻發次數出現在秋季,達1.80×106cells/L。而適宜在較冷環境中引發赤潮的多列擬菱形藻未檢測到,可能與該地區水溫較高有關,在水溫較低的香港海域發現有該藻的存在。

軟骨藻酸對神經膠質細胞HO-1蛋白表達的影響
利用培養的鼠新皮質神經元研究軟骨藻酸的神經毒性行為,發現這3種穀氨酸受體進行性參與神經毒性:(1)間接激活NMDA受體參與神經毒性,(2)由AMPA受體介導的毒性,(3)當受體的脫敏作用被阻斷時,可由KA受體介導神經毒性。軟骨藻酸的劑量-反應關係研究表明,軟骨藻酸可活化KA和AMPA受體,親和力分別約為5和0.07μmol/L。KA受體系統的一個明顯特性是,它在海馬CA3區大量表達。許多動物實驗研究發現它的失控激活可導致癲癇發作和選擇性神經元損傷。體外培養12 d的大腦皮層神經元經軟骨藻酸染毒處理后,有MK-801(一種NMDA受體拮抗劑)和GYKI53655(一種選擇性AMPA受體拮抗劑)存在時,可觀察到小卻顯著的神經元毒性作用,表明此種毒性是由KA受體介導。研究認為可引起明顯神經元毒性劑量水平的軟骨藻酸的作用是通過KA受體的過度激活或興奮毒性刺激而中斷興奮性氨基酸系統,從而損傷學習和記憶功能。Jensen等發現,當GYKI53655存在時可部分消除軟骨藻酸引起的神經毒性,這表明AMPA受體介導部分神經毒性。
MK-801能阻斷部分軟骨藻酸毒性,有人推測NMDA受體間接參與軟骨藻酸的毒性作用,可能是軟骨藻酸作用於神經細胞促進谷氨酸釋放,釋放的谷氨酸繼發激活NMDA受體。在一定的受體位點,閾值水平的軟骨藻酸可能引起內源性興奮氨基酸的過量釋放,例如谷氨酸和天冬氨酸,這些內源性神經遞質可能會增強軟骨藻酸對小鼠腦組織NM-DA受體位點的反應程度。因此當軟骨藻酸的濃度不足以引起神經毒性時,如有NM-DA受體興奮劑的存在,同樣能夠引起神經毒性,因此Antonello等推測,大腦中亞毒性劑量的軟骨藻酸通過持續增強內源性興奮性氨基酸作用於NMDA受體而引起癲癇。在脊椎動物中樞神經系統中,谷氨酸、天冬氨酸是主要的興奮性神經遞質,γ-氨基丁酸是一種重要的抑制性神經遞質。谷氨酸是γ-氨基丁酸的前體。軟骨藻酸可通過促進谷氨酸、天冬氨酸等釋放,抑制谷氨酸攝取及抑制谷氨酸脫羧酶等過程來增加細胞間谷氨酸水平。Dak等曾發現軟骨藻酸可促進大鼠海馬切片釋放谷氨酸,Cunha等報道,3μmol/L軟骨藻酸可抑制24%的γ-氨基丁酸釋放。給妊娠小鼠尾靜脈注射軟骨藻酸,可致後代海馬功能發生重大損害,並伴隨有大腦皮層和海馬γ-氨基丁酸水平下降,而谷氨酸水平升高。釋放出的谷氨酸有助於進一步地興奮毒性損傷,如可加劇軟骨藻酸對分離雞胚視網膜的神經毒性作用。在紋狀體神經元,KA受體的激活可抑制γ-氨基丁酸的作用。谷氨酸介導的損傷主要是經過NMDA受體介導的,軟骨藻酸直接激活AMPA/KA受體釋放谷氨酸和天冬氨酸,谷氨酸和天冬氨酸再激活NMDA受體從而產生興奮毒性。總之,完全程度的神經元衰退是經NMDA受體和非-NMDA受體(AMPA受體和KA受體)協同作用產生的,軟骨藻酸與谷氨酸等內源性氨基酸協同作用以增強其神經毒性。
NMDA受體與突觸的可塑性和學習、記憶密切相關,通過該受體本身、其共軛的離子通道(即NMDA通道)及調節部位三者形成一複合體而發揮功能。每個NMDA受體上含有2個谷氨酸和2個甘氨酸結合識別位點,谷氨酸和甘氨酸均是谷氨酸受體的特異性激活劑。1991年,日本Nakanishi實驗室首次從大鼠腦中成功地克隆出了一種NMDA受體亞基的cDNA(NMDAR1)。此後,人們又相繼克隆出了4個新的NMDA受體亞基的cDNA(NMDAR2 A-D)。NMDAR1可單獨形成功能性純寡聚體NMDA受體,但NMDAR2亞基卻不具備該功能。當NMDAR1與不同的NMDAR2共同表達后,由谷氨酸誘導的電流比對僅NMDAR1獨自表達誘發的電流大數十倍到上百倍,此特性表明NMDA受體可能是由NMDAR1和不同的NMDAR2亞基組成的一個異寡聚體。NMDAR1是由938個氨基酸殘基組成的跨膜蛋白,分子量為105kD,有4個跨膜區。1992年,Novelli等發現,含軟骨藻酸的蚌類對神經培養物的損害與興奮性氨基酸有協同作用,而且他們的資料表明,亞毒作用劑量的軟骨藻酸能促進谷氨酸和精氨酸的興奮毒作用。該協同毒性作用可能是通過降低Mg2+對NMDA受體離子通道的阻礙效應而實現的。Berman等在體外培養的神經元中,利用KA/AMPA及NMDA受體抑製劑,研究了軟骨藻酸對小腦顆粒細胞的毒作用機制。他們的資料顯示,由於NMDA競爭性和非競爭性拮抗劑均能降低軟骨藻酸誘導的內源性興奮性氨基酸的外流,所以NMDA受體不僅能介導大部分軟骨藻酸引起的神經損傷,而且是興奮性氨基酸外流的一個主要部位。這些結果更直接地支持了Novelli等人的觀點。Berman和Heron等還發現,暴露在軟骨藻酸下,內源性腺苷能抑制谷氨酸的釋放。但是,軟骨藻酸和內源性腺苷相互作用的機制還不十分清楚。軟骨藻酸能促進谷氨酸和精氨酸的釋放,而釋放的谷氨酸和精氨酸又可激活NMDA受體,進而促進內源性興奮性氨基酸的釋放。所以軟骨藻酸可協同性地加強谷氨酸/精氨酸介導的神經興奮作用。NMDA受體激活后,細胞外的Ca2+大量進入細胞,細胞內鈣庫貯存的Ca2+大量釋放,引起細胞內Ca2+過負載,造成細胞損傷。高濃度的Ca2+可活化與NMDA受體NR1和NR2B亞基結合的鈣/鈣調素依賴性蛋白激酶Ⅱ(Ca2+/CaM-dependent proteinkinaseⅡ,CaMKⅡ),CaMKⅡ又能磷酸化AMPA受體,進而提高該受體通道的敏感性。
KA/AMPA受體是最重要的離子型非NMDA型興奮性氨基酸受體。AMPA受體家族包括4個結構極為相似的亞基,各亞基的氨基酸序列的同源性高達70%。由於氨基酸殘基疏水性分佈,在靠近羧基端的部分構成4個跨膜區。AMPA、L-谷氨酸及KA均可激活這類離子通道,並有AMPA的高親和力結合位點。天然AMPA受體是一個僅由這4種亞基組成的五聚體,每個單體的分子量為108kD。AMPA受體的4種亞基在第4個跨膜區的上游均含有1個由38個氨基酸殘基組成的特殊區段,該區存在2個結構相似區,分別由受體基因上2個相鄰的外顯子編碼。但各亞基的DNA編碼在翻譯后要經過一些如磷酸化、糖基化及棕櫚酮化等修飾,這些修飾是通道功能重要的調節方式。離子型谷氨酸受體功能的多樣性是通過不同亞基的組裝、選擇性基因接合、轉錄前mRNA的編輯等方式來實現的。特別對於AMPA和KA受體,它們不同於NMDA受體,還要通過RNA編輯進行特殊的修飾。最近發現通道小孔區的氨基酸是編輯酶的作用位點,所以該部位的修飾會影響到離子的通透性。人們繼續研究發現,在AMPA受體GluR5亞基或KA受體GluR5和GluR6亞基的通道小孔區內或附近關鍵位置的RNA編輯,將中性的谷氨酸替換成帶正電荷的精氨酸,谷氨酸和精氨酸的互換會引起Ca2+的內流。成年大鼠的GluR2基因全部經過編輯,而GluR5和GluR6編輯不完全,分別為39%和75%。這種現象可能與編輯有關的腺苷脫氨酶識別和鹼基配對所需的特定內含子序列的位置有關。GluR2的內含子序列在谷氨酸-精氨酸互換點的位置附近,而GluR5和GluR6的內含子序列卻遠離該互換點1900個核苷酸。Seeburg等人研究發現,AMPA受體的電導性和通透性由GluR2決定,這是因為GluR2的第2個跨膜區與其它3個亞基不同,谷氨酸突變為精氨酸。他們還發現,AMPA受體各亞基的基因編碼中,谷氨酸/精氨酸位點均是谷氨酸的編碼。正是由於高度特異性的mRNA編輯才實現了定點突變。結果提示腦細胞具有選擇表達含未編輯GluR2亞基,從而實現選擇對Ca2+通透的AMPA受體數量的能力。
興奮毒素最初通過過度激活谷氨酸受體參與神經元去極化而干擾神經元的調節機制,導致離子滲透壓和電化學的改變,最終引起細胞的死亡。神經毒理學研究表明,軟骨藻酸作用於谷氨酸受體豐富的神經細胞,特別是海馬和錐體細胞,導致大量細胞外Ca2+內流,細胞鈣穩態發生異常改變,信號轉導紊亂;可引起能量代謝障礙,從而引起神經細胞發生退行性變化。
NMDA受體介導細胞內Ca2+濃度的升高是興奮性氨基酸介導的神經元損傷和死亡的一個重要原因。然而非NMDA受體通過細胞膜去極化和激活電壓依賴性Ca2+通道也能引起細胞內Ca2+升高。當神經元去極化,結合軟骨藻酸的AMPA受體打開了配體門控離子通道允許Na+內流,K+外流,氯化物隨Na+進入細胞,隨後在滲透壓的作用下水也內流;這樣就產生了長時間的突觸后腫脹,繼之造成細胞內鈣離子超載並發展為持久的神經元損傷。而Bureau等發現星形膠質細胞的AMPA受體對Ca2+有滲透性,Ca2+可通過AMPA受體通道直接進入細胞。KA受體的過度興奮引起癲癇活動的擴散,通過內源性谷氨酸致使其它谷氨酸受體包括NMDA受體激活,可引起大量細胞去極化,並激活電壓依賴性Ca2+通道促使細胞內Ca2+升高。長時間的細胞內遊離Ca2+的升高是有毒的,並且最終引起神經元變性。NMDA受體激活后細胞外的Ca2+大量進入細胞,細胞內鈣庫儲存的Ca2+大量釋放,引起細胞內Ca2+超載,造成細胞損傷。而AMPA/KA受體介導的去極化也可能通過降低Mg2+對NMDA受體離子通道的阻礙效應而提高胞質內遊離Ca2+濃度。同時,由軟骨藻酸刺激釋放的內源性谷氨酸也與NMDA受體相互作用促使突觸后膜去極化,引起電壓依賴性Ca2+通道和興奮劑激活的Ca2+通道的開放,促使Ca2+大量內流,造成細胞內鈣積聚,結果引起細胞壞死。NMDA受體的電壓依賴本質及其與第二信使鈣離子的作用相結合,在學習記憶機制中起主要作用。
局部大腦功能活動與局部大腦葡萄糖代謝率相關。在軟骨藻酸中毒人群中,用正電子發射體層攝影術觀察到在海馬和杏仁核的葡萄糖代謝率降低。在一定的病理條件下,神經元不具有足夠的代謝能量來維持正常的靜息膜電化學梯度。當神經元胞體表面或樹突附近谷氨酸過量時,產生神經元去極化和胞漿膜滲透性持續性升高,隨後細胞離子組成不平衡激活了利用能量維持功能的膜泵。由於胞漿內遊離鈣過多,隨後進入和積聚在線粒體內,損傷氧化磷酸化,造成ATP合成不足;另一方面由於肌纖維、肌漿網和線粒體中鈣依賴性ATP酶的超常活動,使ATP消耗增多。而神經元的持續激活將導致儲存能量的不可逆衰竭和神經元細胞不能維持離子平衡。這必然導致細胞功能和結構的破壞,通過細胞離子成分的改變有可能伴隨著水運動的改變,導致胞漿空泡的形成,細胞腫脹,最後組織壞死。另外,由於缺少ATP而使Ca2+-ATP酶活性降低,這似乎有助於促進軟骨藻酸升高細胞內Ca2+水平。由興奮性氨基酸引起的神經毒性在低能量的神經元大大增加,因此軟骨藻酸對年紀大的人的危害更嚴重。
軟骨藻酸能促進谷氨酸、天冬氨酸釋放,抑制谷氨酸攝取及抑制谷氨酸脫羧酶增加突觸間隙谷氨酸濃度。谷氨酸、天冬氨酸是最為重要的興奮性氨基酸遞質。中樞的谷氨酸並非來自外周,主要是在中樞以谷氨醯胺為前體在谷氨醯胺酶作用下自身合成。同時激活代謝型受體,導致細胞內三磷酸肌醇和甘油三脂形成增加,胞內儲存的Ca2+釋放,進一步提高胞內Ca2+水平,導致鈣穩態失衡,從而引起神經興奮和(或)興奮性毒性。實驗中,細胞在中、高劑量的軟骨藻酸作用2h后,釋放出胞外的天冬氨酸和谷氨酸顯著升高,這與Ross等報道的軟骨藻酸引起大鼠膠質細胞興奮性氨基酸升高相一致,提示軟骨藻酸確可引起膠質細胞興奮性氨基酸升高。各劑量軟骨藻酸均使胞內[Ca2+]升高,且有明顯的劑量-效應關係,提示軟骨藻酸對[Ca2+]變化的影響比對氨基酸釋放影響更加敏感。興奮性氨基酸濃度與胞內[Ca2+]有高度的相關性,胞內[Ca2+]蓄積可通過CaM或直接作用於細胞膜,最終導致神經細胞膜的通透性增高,水分增多,細胞腫脹損傷。同時,胞內[Ca2+]濃度的升高和細胞的腫脹有直接促進興奮性氨基酸的釋放,從而加重興奮性神經毒性。
抑制性氨基酸表現為抑制效應,與興奮性氨基酸共同作用調節神經系統的興奮性和興奮毒性。γ-氨基丁酸(GABA)是中樞神經系統主要的抑制性神經遞質,通過谷氨酸脫羧酶(glutamic acid devarboxylase,GAD)催化谷氨酸脫羧而成,主要通過線粒體的GABA-α-谷氨酸轉移酶分解代謝再生成谷氨酸。實驗中,各劑量組GABA均比對照組顯著下降,呈劑量-效應關係,與Sobotka等報道的軟骨藻酸抑制GABA釋放相符,可能是由於胞內GABA的前體---谷氨酸濃度降低所致。甘氨酸變化不顯著,僅高劑量組較對照組明顯升高。研究中發現甘氨酸與興奮性氨基酸遞質升高的同步性揭示甘氨酸在軟骨藻酸的興奮性神經毒性中可能也發揮著一定的作用。另外,研究提示在軟骨藻酸的興奮性神經毒性中,興奮性氨基酸發揮主要作用,抑制性氨基酸起協同作用。
該法是根據美國公職化學家協會(AOAC)所標定的麻痹性貝中毒(PSP)的小鼠生物分析法加以變化的,是最早用於DA檢測的一種經典方法。其原理是將毒素注射入小白鼠體內,根據注射后小鼠的存活時間和中毒癥狀來評價毒性,一般用半致死劑量表示。1987 年該方法首次應用於加拿大DA 中毒事件中DA 的測定,結果顯示該方法適於檢測貝類組織中高濃度的DA(高於30μg/g),DA 濃度較低時無法檢測(低於20μg/g)。
該法更加註重研究DA的毒性效應和毒理研究。這是一般其他方法無法取代的,而且採用小白鼠生物檢測法可以檢測一些未知的毒性物質,通過半數致死劑量來評價該物質的毒性效應,因此小白鼠生物檢測法在各個國家仍有很廣泛的應用。但是,該方法的檢出限較高,適用於檢測貝類組織中高濃度的DA(300-1000μg/g),不適用於DA含量低於20μg/g組織的檢測。
小白鼠生物檢測法方法檢測周期大於24h,特異性較差,干擾因素較多,樣品處理為粗略提取,樣品批次單一。小白鼠生物檢測法檢測DA是最早使用的一種方法,但是其試驗結果受外部影響因素較為複雜,試驗結果與小白鼠的品系和試驗者本身操作有很大關係,並且小白鼠生物檢測法一般要有至少1d的檢測周期,該方法的檢出限較低。此外,小白鼠生物法僅能測定毒性的大小,無法確定毒素的組成和各成分的含量。小白鼠生物檢測法最大的局限性是小鼠本身造成結果的高變異性和低敏感性,而且對鼠種的要求較高。
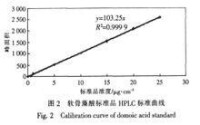
軟骨藻酸HPLC曲線
軟骨藻酸HPLC曲線參考文獻。
DA-HPLC檢測DA的方法主要有兩大類,一種方法是貝類樣品被提取后,經濃縮純化後進行HPLC配紫外檢測器(HPLC-UVD)分析。由於DA在242nm的紫外光譜區有最大吸收峰,因此反相高效液相色譜(RP-HPLC)-UV可有效而快速地對樣品進行分離和檢測,其基本原理是使用酸性流動相以抑制羧基的電離作用,並選用C18鍵合硅膠柱分離DA及其衍生物。HPLC-UVD法特別適用於分析貝類組織內的DA含量,其檢測限在10-80ng/ml之間,且依提取和提純方法的不同而異,如粗提取物未經提純,其檢測限約為1μg/g,適用於毒素含量超過20μg/g的檢測。水中DA的檢測與樣品量及藻類的含量和種類等因素有關。另一種方法是針對海水和硅藻中DA進行的甲氧基芴甲酸(9-fluorenylmethyloxycarbonyl,FMOC)衍生化/HPLC配熒光檢測器(HPLC-FLD)法,該法在DA分子上引進了熒光基團而使FLD檢測的靈敏度提高,對海水中DA的檢測限為1.5pg/ml。Hummert等提出了柱切換系統,它能有效降低提取物中的干擾,直接分析粗提的樣品,省去了複雜的樣品預處理過程。流動相的配比影響DA的保留時間,隨著流動相中乙腈比例的減小,DA保留時間增加,DA色譜峰對稱性增強,DA與DAC′5非對映異構體分離度增大。中國報道用RP-HPLC-UVD測定貝類中DA,經甲醇:水(1:1)溶液提取,強陰離子(stronganion exchange,SAX)固相萃取柱凈化,C18反相色譜柱分離,流動相為乙腈:水(13:87),242nm波長下檢測,最低檢出限為0.2μg/g,在1.0-25.0μg/ml範圍內有良好的線性關係,平均回收率為(97.23±2.43)%。流動相採用甲醇:水(22:78,pH2),最低檢出限為10ng(1.0μg/ml),檢測範圍50-250ng(5-25μg/ml),DA的回收率可達(99.9±2.4)%。另有報道流動相為甲醇:乙腈:三氟乙酸(80:20:0.1),最低檢出限為10ng(1.0μg/ml),在1-40μg/ml範圍內線性關係良好。
高效液相色譜-紫外檢測法(HPLC-UV)特別適用於貝類組織中DA含量的測定,尤其適用於毒素含量超過20μg/g的情況。Lawrence等人建立[以及Quilliam等改進的HPLC-UV法採用了反相C18柱在242nm處檢測DA,該方法已被英國、愛爾蘭等國定為標準方法。後來,又有一些研究人員對此方法進行了改進。López-Rivera等建立了一種不需要固相萃取預處理檢測DA的HPLC-UV法,通過等梯度淋洗以及控制流動相pH來分離化合物,結果表明,最佳pH為2.5,方法檢測限為25ng/mL。該方法快速、靈敏,能成功檢測大批量貝類樣品。HPLC-UV法已經被很多研究人員用來檢測貝類等動物組織中的DA含量。Costa等用HPLC-UV法檢測了葡萄牙海岸的兩種章魚E.cirrhosa和E.moschata體內DA的含量,結果表明,DA含量超過了100μg/g,其中,E.moschata體內毒素含量更高,表明DA可能通過食物鏈由這種章魚體內進入更高級消費者如人類體內,潛在危險性更大。中國對DA的檢測大多採用HPLC-UV法,張一兵等在借鑒國外方法的基礎上應用高效液相色譜-紫外可見檢測法(HPLC-UVD)進一步優化了DA的高效液相色譜-紫外檢測法,選定pH2的甲醇/水=22∶78(V/V)為流動相,LC-SAX作為濃縮純化柱,檢出限為10ng,回收率達到92%-94%。陳西平等採用該方法檢測了部分水及水生動物中DA的含量,回收率達到70%-93%,檢出限為10ng。結果表明,雖然在海水及地面淡水中未檢出DA,但部分海洋貝殼動物體內有檢出。因此,隨著海洋污染的加劇,現階段中國的DA污染問題不容忽視,應加強對其污染狀況的監測。
高效液相色譜-質譜(HPLC-MS)聯用技術可以不使用衍生試劑和毒素標準品,相比其他檢測器,具有很大的優勢。然而本方法對設備條件要求較高。Lawrence等建立的高效液相色譜-電噴霧離子阱質譜法(HPLC/ESI-MS)法是一種高靈敏度、高選擇性的方法,被應用到了環境中各種介質的檢測中,並被後人不斷改進,但是這種方法對樣品預處理的要求較高。宋琍琍等參考了這種方法來測定DA殘留。樣品經50%甲醇提取,LC-SAX 柱凈化,然後選用電噴霧離子源進行測定分析,該方法的定量限為0.02μg/g。Pardo等採用加壓濕法萃取(PLE)來提取DA,然後用液相色譜-電噴霧電離雙質譜(HPLC-ESI-MS-MS)法來檢測DA,電離源採用電噴霧可以提高分析信號,方法經過優化后回收率在81%至95%之間,他們用此方法成功檢測了46個西班牙超市裡的貝類樣品,表明該方法能進行大批量檢測。Wang 等採用LC-MS 對水樣和浮游植物中的DA 進行了定性和定量分析,樣品預處理採用C18柱進行固相萃取,結果表明,酸性條件有利於在C18 柱上保留親水性的DA,加標水樣的回收率超過了90%,浮游植物樣品的回收率則接近98%,方法檢測限為0.03ng/mL。Iglesia等也採用LC-MS法檢測了海水中的DA及它的異構體,但樣品預處理採用的是固相萃取盤,該方法的檢測限為0.02ng/mL,回收率為92.1%-110.6%,用此方法能檢測海水中的痕量DA。Vale等利用HPLC-MS的方法,以1+1的甲醇溶液作為洗脫液,對葡萄牙魚類和貝類體內的軟骨藻酸含量進行測定,結果表明在春季樣品中DA含量最高;在分析的眾多樣品中,蛤仔和鳥蛤被軟骨藻酸污染最為嚴重,紫貽貝和牡蠣等物種受污染較輕。陳燕青等利用HPLC-MS法測定蝦夷扇貝和長牡蠣中軟骨藻酸的殘留,樣品經濃度50%甲醇提取,LC-SAX柱凈化,3ml0.1mol/L甲酸溶液洗脫,電噴霧離子源(ESI),在正離子、多反應監測方式(MRM)模式下進行定性與定量,該方法的檢出限達0.01μg/g,線性範圍為0.02-10.00μg/g。
高效液相色譜-熒光檢測法是使用衍生劑將DA衍生成含熒光基團的物質,用熒光檢測器進行檢測,在DA分子上引入熒光基團可使方法的靈敏度提高。檢出限可達到pg/ml。Maroulisb等利用柱后衍生法測定DA,加入4-氯-7-硝基-2,1,3-苯並氧雜惡二唑(NBD-Cl)與DA上的氨基反應生成熒光基團,提高了熒光檢測檢測器(FLD)的靈敏度,熒光檢測器激發波長設為469和529nm,檢出限低至25μg/kg。James等使用NBD-F來衍生化DA,用等梯度淋洗程序分離化合物,激發波長為470nm,發射波長為530nm,方法檢測限高於1ng/mL,貝類組織中DA的回收率>95%,當用強陰離子交換SPE柱萃取DA時,檢測限達到6ngDA/g貝類組織。Chan等參考了這種方法來檢測DA,但他們改進了樣品預處理方法,即在SPE柱中加入TiO2,可以更好地吸附DA,並探討了最佳pH,結果表明,吸附的最佳pH為4,解吸時pH則為11,吸附平衡時間為2小時,最佳吸附劑裝載量為0.67mgTiO2/ngDA。Maroulis等採用NBD-Cl柱后衍生DA進行檢測,成功測得了貝類組織中DA含量為75μg/kg的樣品,該方法檢測限低至25ppb,能實現全自動化分析,對樣品預處理要求低,干擾少。由於大多數衍生試劑價格昂貴、不穩定,且其分解產物可能出現在色譜圖中,同時衍生試劑的變質也可能導致毒素的不完全衍生化,因此熒光檢測法的實際應用很少。
免疫分析法(ELISA)是基於抗體與抗原或半抗原之間的高選擇性反應而建立起來的一種生物化學分析法,即根據抗原-抗體反應,採用特異性貝類毒素的抗體來檢測DA。免疫分析法主要包括酶聯免疫法、放射性免疫法、熒光免疫法等,具有方便、快速的特點,但缺點是費用較高,而且各毒素之間的交叉反應低,不能全面體現出樣品的毒性。檢測DA所採用的免疫分析方法主要是酶聯免疫吸附法(ELISA)。ELISA是利用抗原-抗體反應的特異性與酶催化作用的高效性相結合,通過酶作用於底物后的顯色反應判定結果,是應用最廣泛的免疫學檢測技術,由於其實驗結果可用目測,也可用酶標儀測定光密度值以反映抗原含量,所以有易定性定量的雙重優點。Engvall和Perlmann於1971年最先應用該法進行IgG定量測定,並命名為Enzyme linked immunosor-bentassay(ELISA)。由於ELISA方法靈敏,特異性強不需要特殊設備,被廣泛應用於各種抗原和抗體測定。將ELISA應用於DA的檢測中,利用原-抗體反應,採用特異性貝類毒素的抗體來檢測DA。
Yu等採用匙孔血藍蛋白(KLH)和小牛血清蛋白(BSA)為載體,進行直接酶聯免疫測定,結果表明,藍貽貝樣品的檢測限<25ng/g,回收率為81.1%,Yu等還將實驗結果用HPLC方法進行了驗證,最終發現15種被污染的貝類樣品中有10種樣品的DA含量<50ng/g。Maucher等結合ELISA技術使用血液採集卡來提取小鼠血液中的DA,這種方法可以避免DA在血液中被清除。一般說來,在4小時內,99%的DA會從血液中被清除,但使用該方法后,DA在24小時內仍然能被檢測到。該方法靈敏度高,用血液採集卡提取樣品方便,因此可用此方法來監測海洋哺乳動物體內的DA含量。Tsao等採用單克隆抗體ELISA檢測法檢測DA,並建立了基於單克隆抗體的膠體金免疫條檢測法,免疫條檢測法的檢測限為5ng/mL,在十分鐘內就可完成檢測。Tsao等的實驗結果表明,用免疫條檢測得到的結果與用ELISA方法得到的結果有很好的吻合性,可以用來快速檢測貝類樣品中的DA。另外,Shaw等建立了基於基因工程單鏈抗體(scFv)競爭ELISA檢測方法,相比傳統方法,這種方法具有很多優勢,基因工程單鏈抗體(scFv)可以在大腸桿菌中快速、大量地得到表達,且容易和其他小分子融合,形成融合蛋白以用於檢測或其它用途。用該方法檢測天然扇貝組織得到的數據與標準HPLC方法檢測的結果基本相符。大多數ELISA法採用的都是直接法,而許道艷等則以DA-OVA為包被抗原,利用抗原抗體反應,建立了間接競爭酶聯免疫吸附技術分析檢測海水樣品和海洋貝類中DA的方法。結果表明,該方法最低檢出限為10μg/L,海水樣品平均回收率為102.2%,貝類樣品平均回收率為111.5%。ELISA方法能夠進行批量檢測,但特異性較差,因而主要用於DA的初步篩選。由於DA標準品昂貴,且DA分子量太小,因此製備免疫抗原較困難,從而限制了免疫方法在DA分析中的應用。

軟骨藻酸毛細管電泳色譜
液質聯用(LC-MS/MS)技術幾乎可以分析所有的貝毒。液質聯用依賴於高效液相色譜把貝毒成分通過離子化帶到質譜中。電噴霧電離源(ESI)和大氣壓化學電離源(APCI),這兩種技術都已經應用於貝毒分析。由於食品(動物組織)的背景非常複雜,樣品間差異又相當大,採用LC-MS(MS/MS)技術測定食品中貝類毒素的關鍵是樣品的前處理。國外文獻方法多採用甲醇-水溶液提取,然後固相萃取法進行凈化與濃縮,最常用的固相萃取柱有Cl8烷基鍵合硅膠柱,CN柱和離子交換柱,最常用的是陰離子交換柱,非鍵合硅膠柱也有很好的純化和萃取效果。此外還有溶劑提取方法、液相色譜製備法和超臨界流體萃取法等。食品(水產品)中的貝類毒素殘留量甚微,一般為μg/kg水平。2005年方曉明等利用液相色譜四極桿-飛行時間質譜(LC/Q-TOFMS)聯用技術對貝類樣品的失憶性貝毒進行了研究。樣品經甲醇/水(體積比為1:1)提取,經SAX固相萃取柱凈化后,用Zorbax Eclipse XDB-18柱(250mm×2.1mm,3.5μm)色譜分離,以乙腈/水含0.05%甲酸水溶液作流動相,梯度淋洗,流速為0.20mL/min。Q-TOFMS選擇電噴霧正離子模式,以[M+H]+作為前體離子進行TOFMS掃描。結果表明:樣品加標的平均回收率為87.8%-94.6%,相對標準偏差為8.4%-11.9%,檢測低限(LOQ)為0.1μg/g。由於Q-TOFMS具有高解析度,能進行精確測定而不降低靈敏度,因此本方法作為對貝類樣品中失憶性貝毒的確證分析方法具有高靈敏度和選擇性。2003年HollandPT等用LC-MS/MS的方法測定了貝類中軟骨藻酸和其異構體的含量,以甲醇/水(體積比為1:1)作為提取劑,以乙腈/水(體積比為1:1)作為稀釋劑,經SPE固相萃取后,用C18柱色譜分離。選用了4種不同的貝類,當軟骨藻酸在樣品中的含量為1μg/g貝-2μg/g貝時,軟骨藻酸的回收率為93%(RSD大於7%)。2007年PardoO等用甲醇/乙腈(體積比為9:1)作為提取劑,經PLE萃取后,通過電噴霧電離(ESI)界面與質譜聯用(ESI-MS-MS)分析測定軟骨藻酸。使軟骨藻酸在0.05-5μg/mL範圍內具有良好的線性關係,相關係數r=0.999;當方法檢出限為0.2μg/g,回收率為81%;當方法檢出限為0.5μg/g,回收率為95%。此方法成功應用於46個貝類樣品的檢測中。2007年WangZH採用離子碎片的方式,使用多反應器,以甲醇/水(體積比為1:1)作為提取劑,以乙腈/水(體積比為1:1)作為稀釋劑,用4mL0.15%甲酸洗脫,經SPE固相萃取后,用C18柱色譜分離,當檢測濃度為30pg/mL,使得回收率從90%提高到98%,建立了一種定量和定性的方法。
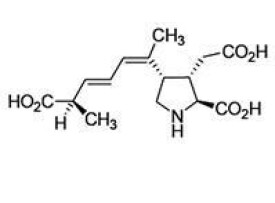
基本信息
- 中文名
- 軟骨藻酸
- 外文名
- domoic acid
- 別名
- DA
- 化學式
- C15H21NO6
- 熔點
- 223-224℃
- 摩爾質量
- 311.33
- 外觀
- 白色固體粉末
- 主要危害
- 神經毒素
- CAS號
- 14277-97-5
- 毒性
- LD50約10mg/kg