ABO血型系統
ABO血型系統
ABO血型系統
血型實質上是不同的紅細胞表面抗原。紅細胞質膜上的鞘糖脂是AB0血型系統的血型抗原,血型免疫活性特異性的分子基礎是糖鏈的糖基組成。1960年,瓦特金斯(A. Watkins)確定了ABO抗原是糖類,並測定了其結構。A、B、O三種血型抗原的糖鏈結構基本相同,只是糖鏈末端的糖基有所不同。A型血的糖鏈末端為N-乙醯半乳糖胺;B型血為半乳糖;AB型兩種糖基都有,O型血則缺少這兩種糖基。
在ABO抗原的生物合成中三個等位基因ABO及H控制著A、B抗原的形成。ABO抗原的前體是H抗原;A基因編碼一種叫N-乙醯半乳糖胺轉移酶的蛋白質(A 酶),能把H抗原轉化成A抗原;B基因編碼一種叫半乳糖轉移酶的蛋白質(B酶),能把H抗原轉化成B抗原;O基因不能編碼有活性的酶,而只有H抗原。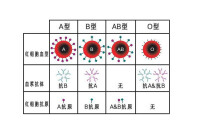
ABO血型系統
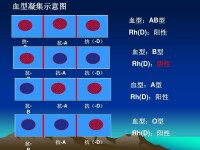
輸血時若血型不合會使輸入的紅細胞發生凝集,引起血管阻塞和血管內大量溶血,造成嚴重後果。所以在輸血前必須作血型鑒定。正常情況下只有ABO血型相同者可以相互輸血。在缺乏同型血源的緊急情況下,因O型紅細胞無凝集原,不會被凝集,可輸給任何其他血型的人。AB型的人,血清中無凝集素,可接受任何型的紅細胞。但是異型輸血輸入量大時,輸入血中的凝集素未能被高度稀釋,有可能使受血者的紅細胞凝集。所以大量輸血時仍應採用同型血。臨床上在輸血前除鑒定ABO血型外,還根據凝集反應原理,將供血者和受血者的血液作交叉配血實驗,在體外確證兩者血液相混不發生凝集,方可進行輸血以確保安全。
近年來發現存在於紅細胞上的凝集原(即血型抗原)也存在於其他血細胞和一般組織細胞。所有細胞表面血型抗原的特異性可作為機體免疫系統鑒別自身和異物的標誌。因此,在臨床實踐中血型鑒定也是組織器官移植成敗的關鍵。人類血型有遺傳特性,決定血型的血型抗原即凝集原ABOA和B及其前身物H,分別受ABO三個等位基因控制。A、B基因為顯性基因,O(H)基因為隱性基因。它們的遺傳規律所顯示的父、母血型與子代血型間的關係,在法醫學上可作為否定親子關係的依據,若再配合其他血型系統的測定,則可判斷親子關係。
| 父母血型 | 子女可能有血型及比例 | 子女不可能有血型 |
| O、O | O | A、B、AB |
| O、A | O、A (1:3) | B、AB |
| O、B | O、B (1:3) | A、AB |
| O、AB | A、B (1:1) | O、AB |
| A、A | O、A (1:15) | B、AB |
| A、B | A、B、AB、O (3:3:9:1) | — |
| A、AB | A、B、AB (4:1:3) | O |
| B、B | O、B(1:15) | A、AB |
| B、AB | A、B、AB(1:4:3) | O |
| AB、AB | A、B、AB(1:1:2) | O |
血型不僅在輸血上有重要意義,而且在人種學、遺傳學、法醫學、移植免疫、疾病抵抗力(或易感性)等方面都有應用價值,在輸血前,一定要檢查病人(受血者)和輸血人(供血者)的血型,並且要進行交叉配血試驗。在臨床醫學中,除輸血、移植免疫外,對新生兒溶血病、自身免疫性溶血性貧血特異性抗體的檢查,也都需要血型知識和有關技術。
血型
血型(blood groups;blood types)是以血液抗原形式表現出來的一種遺傳性狀。
狹義地講,血型專指紅細胞抗原在個體間的差異;但現已知道除紅細胞外,在白細胞、血小板乃至某些血漿蛋白,個體之間也存在著抗原差異。因此,廣義的血型應包括血液各成分的抗原在個體間出現的差異。通常人們對血型的了解往往僅局限於ABO血型以及輸血問題等方面,實際上,血型在人類學、遺傳學、法醫學、臨床醫學等學科都有廣泛的實用價值,因此具有著重要的理論和實踐意義,同時,動物血型的發現也為血型研究提供了新的問題和研究方向。血型一般常分A、B、AB和O四種,另外還有RH、MNS、P等極為稀少的10餘種血型系統。其中,AB型可以接受任何血型的血液輸入,因此被稱作萬能受血者,O型可以輸出給任何血型的人體內,因此被稱作萬能輸血者、異能血者、實際上,不同血型之間的輸送,一般只能小量的輸送,不能大量。要大量輸血的話,最好還是相同血型之間為好。
科學史記載:在17世紀80年代的英國,有位醫生曾經給一個生命垂危的年輕人輸羊血,奇迹般的挽救了他的生命。其他醫生紛紛效仿,結果造成大量受血者死亡。
直到20世紀初,我們才打開了科學輸血的大門。人類最早認識的血型系統是ABO血型系統。1900年,奧地利維也納大學病理研究所的研究員卡爾·蘭德施泰納發現:健康人的血清對不同人類個體的紅細胞有凝聚作用。如果把取自不同人的血清和紅細胞成對混合,可以分為A、B、C(后改稱O)三個組。後來,他的學生Decastello和Sturli又發現了第四組,即AB組。
紅細胞血型是1900年由奧地利的K.蘭德施泰納發現的。他把每個人的紅細胞分別與別人的血清交叉混合后,發現有的血液之間發生凝集反應,有的則不發生。他認為凡是凝集者,紅細胞上有一種抗原,血清中有一種抗體。如抗原與抗體有相對應的特異關係,便發生凝集反應。如紅細胞上有A抗原,血清中有A抗體,便會發生凝集。如果紅細胞缺乏某一種抗原,或血清中缺乏與之對應的抗體,就不發生凝集。根據這個原理他發現了人的ABO血型。後來他又把不同人的紅細胞分別注射到家兔體內,在家兔血清中產生了3種免疫性抗體,分別叫做M抗體、N抗體及P抗體。用這3種抗體,又可確定紅細胞上3種新的抗原。這些新的抗原與ABO血型無關,是獨立遺傳的,是另外的血型系統。而且M、N與P也不是一個系統。控制不同血型系統的血型基因在不同的染色體上,即使在一個染色體上,兩個系統的基因位點也相距甚遠,不是連鎖關係,因此是獨立遺傳的。
Rh是恆河猴(Rhesus Macacus)外文名稱的頭兩個字母。蘭德斯坦納等科學家在1940年做動物實驗時,發現恆河猴和多數人體內的紅細胞上存在Rh血型的抗原物質,故而命名的。凡是人體血液紅細胞上有Rh抗原(又稱D抗原)的,稱為Rh陽性。這樣就使已發現的紅細胞A、B、O及AB四種主要血型的人,又都分別一分為二地被劃分為Rh陽性和陰性兩種。隨著對Rh血型的不斷研究,認為Rh血型系統可能是紅細胞血型中最為複雜的一個血型系。Rh血型的發現,對更加科學地指導輸血工作和進一步提高新生兒溶血病的實驗診斷和維護母嬰健康,都有非常重要的作用。根據有關資料介紹,Rh陽性血型在我國漢族及大多數民族人中約佔99.7%,個別少數民族約為90%。在國外的一些民族中,Rh陽性血型的人約為85%,其中在歐美白種人中,Rh陰性血型人約佔15%
在我國,RH陰性血型只佔千分之三到四。RH陰性A型、B型、O型、AB型的比例是3:3:3:1。
有些血型抗體是不完全抗體,與相應的抗原細胞結合后看不出凝集現象,血清中有抗體但不容易發現。1945年抗人球蛋白試驗應用到血型檢查中來,這種試驗就可檢查不完全抗體,從此,許多血型抗原陸續被人發現。每當發現一個新抗原后就要確定這一抗原與已經發現的血型是什麼關係,這樣在人的紅細胞上便確定了若干血型系統。此外,還有一些抗原,或因其在群體中出現的頻率太高,或因其在群體中分佈的頻率太低,對它們無法進行遺傳學分析。在沒有弄清它們的遺傳關係以前,暫且把這些抗原分別叫做高頻率抗原及低頻率抗原,對於它們的歸屬有待進一步確定。
紅細胞膜上另一類血型抗原叫MN抗原,即紅細胞膜上的血型糖蛋白A。它在SOS凝膠電泳譜上顯示兩條區帶,即PAS-1和PAS-2,血型糖蛋白A是兩者的二聚物。已知血型糖蛋白A由131個氨基酸組成,其一級結構已測定(圖2)。血型糖蛋白A的肽鏈呈三節式結構,中間第73~92號氨基酸為疏水性肽鏈,可橫穿膜脂層;N端肽鏈位於膜外側,與血型活性有關,在這段肽鏈上分佈有15條O-糖苷鍵型糖鏈和1條N-糖苷鍵型糖鏈,糖鏈中唾液酸占紅細胞膜上全部唾液酸的一半以上;C端肽鏈位於膜內側,含較多酸性氨基酸。
MN抗原由M抗原和N抗原兩部分組成,如果用神經氨酸酶將M抗原切去1個唾液酸(N-乙醯神經氨酸),則為N抗原,如再切去一個唾液酸則抗原性完全失去。MN抗原的抗原性還和肽鏈上的氨基有關,若將氨基用乙醯基保護后即失去抗原性。隨著S和s兩個抗原的發現,此血型系統一般稱為MNS血型系統。
HLA血型系統是人類白細胞抗原中最重要的一類。與紅細胞血型相比,人們對白細胞抗原的了解較晚,人體第一個白細胞抗原Mac是1958年法國科學家J.多塞發現的。HLA是人體白細胞抗原的英文縮寫,已發現HLA抗原有144種以上,這些抗原分為A、B、C、D、DR、DQ和DP7個系列,而且HLA在其他細胞表面上也存在。
HLA抗原是一種糖蛋白(含糖為9%),其分子結構與免疫球蛋白極相似(圖3)。HLA分子由4條肽鏈組成(含2條輕鏈和2條重鏈),重鏈上連接2條糖鏈。HLA分子部分鑲嵌在細胞膜的雙脂層中,其插入膜的部分相當於免疫球蛋白IgG的Fc區段,輕鏈為β-微球蛋白。由於分子結構上的相似,故HLA與有保衛功能的免疫防禦系統密切相關。
此外,HLA和紅細胞血型一樣都受遺傳規律的控制。決定HLA型的基因在第6對染色體上。每個人分別可從父母獲得一套染色體,所以一個人可以同時查出A、B、C、D和DR5個系列中的5~10種白細胞型,因此表現出來的各種白細胞型有上億種之多。在無血緣關係的人間找出HLA相同的兩個是很困難的。但同胞兄弟姊妹之間總是有1/4機會HLA完全相同或完全不同。因此法醫鑒定親緣關係時,HLA測定是最有力的工具。
紅細胞膜中夾雜著3種蛋白質:糖蛋白、簡單蛋白及膜收縮蛋白。紅細胞抗原有些突出在細胞表面,好像伸出在地面上的樹枝,如ABH抗原;有些鑲嵌在細胞膜內,如Rh抗原。抗原與抗體發生特異反應的部分,叫做抗原決定簇。血型抗原決定簇的化學組成,有的已經清楚,但大部分不清楚。有些血型在體液中存在可溶性抗原,叫做血型物質。從人體分離出來的ABH及Lewis血型物質是糖蛋白,即在肽鏈的骨架上連接著一些糖的側鏈,這些糖鏈便是特異性決定簇。ABH及Lewis血型物質的特異性決定簇很相似,只是在糖鏈上個別糖的種類或同一種糖由於存在位置不同,就顯出不同的特異性。比如A與B的抗原特異性,只是在糖鏈上有一個糖不相同,便顯示出不同的特異性。A抗原決定簇在糖鏈的終末端是一個N-乙醯半乳糖胺,而B抗原決定簇在糖鏈的終末端卻是一個D-半乳糖。
紅細胞上的ABH抗原決定簇,雖與體液中的抗原決定簇糖鏈結構相同,但連接的骨架不同。紅細胞上的糖鏈是通過神經鞘氨醇與脂肪酸結合在一起,而不是與蛋白質結合在一起,所以紅細胞上的ABH抗原是糖脂而不是糖蛋白。
MN·P及I血型的抗原決定簇也是碳水化合物。Rh抗原的決定簇可能是蛋白質,因為紅細胞經硫氫化物、脲素及蛋白酶等物處理后,Rh活性即行消失。
有一些血型抗體,如抗IH,抗IA,抗IB,抗IP1等,只與帶有I抗原及另外一個抗原的細胞發生反應,而不與其中只有一個抗原的細胞發生反應。說明這些抗原為複合抗原,在一個分子上具有兩種特異性。
Lewis血型抗原實際上是血漿中的抗原,紅細胞上的Lewis抗原是從血漿中吸附來的。I抗原在分泌液中雖有可溶性抗原,但不存在於血漿中。另外有些血型是在血漿中存在可溶性抗原,分泌液中卻不存在。Bg抗原實際是白細胞的抗原,可能從白細胞脫落到血漿中,再從血漿中吸附到紅細胞上,表現為紅細胞的抗原。Chido血型及Rodger血型的抗原與血漿中的補體第四成分(C4)有關。用電泳方法分析人的C4,可以見到3種類型:泳動快的(F);泳動慢的(S);快慢兩種成份都有的(FS)。血漿中只有F成份的人,紅細胞上有Rodger抗原。只有S成份的人,紅細胞上有Chido抗原。兩種成份全有的人,紅細胞上也同時具有Chido及Rodger兩種抗原。
各種血型抗原在紅細胞上的分佈是不同的,有的密集,有的疏鬆。抗原數目的多少決定了抗原的強弱。用放射性碘標記的兔抗A及抗B血清,檢查人的紅細胞,根據每個細胞上的放射性強度,可以推算出每個紅細胞上的抗原數目。
基本信息
- 中文名
- ABO血型系統
- 外文名
- ABO blood group system
- 定義
- 根據紅細胞表面有無特異性抗原(凝集原)A和B來劃分的血液類型系統
- 其他血型系統
- Rh血型系統 MN血型 HLA血型系統
- 分子基礎
- 不同的紅細胞表面抗原
- 輸血
- 所以在輸血前必須作血型鑒定
- 概述
- 根據紅細胞表面有無特異性抗原