脂肪酸
由碳、氫、氧三種元素組成的一類化合物
脂肪酸(fatty acid),是指一端含有一個羧基的長的脂肪族碳氫鏈,是有機物,直鏈飽和脂肪酸的通式是C(n)H(2n+1)COOH,低級的脂肪酸是無色液體,有刺激性氣味,高級的脂肪酸是蠟狀固體,無可明顯嗅到的氣味。
脂肪酸是最簡單的一種脂,它是許多更複雜的脂的組成成分。脂肪酸在有充足氧供給的情況下,可氧化分解為CO2和H2O,釋放大量能量,因此脂肪酸是機體主要能量來源之一。
脂肪酸主要用於製造日用化妝品、洗滌劑、工業脂肪酸鹽、塗料、油漆、橡膠、肥皂等。
脂肪酸是由碳、氫、氧三種元素組成的一類化合物,是中性脂肪、磷脂和糖脂的主要成分。脂肪酸代謝脂肪酸根據碳鏈長度的不同又可將其分為:短鏈脂肪酸,其碳鏈上的碳原子數小於6,也稱作揮發性脂肪酸;中鏈脂肪酸,指碳鏈上碳原子數為6-12的脂肪酸,主要成分是辛酸(C8)和癸酸(C10);長鏈脂肪酸,其碳鏈上碳原子數大於12。一般食物所含的大多是長鏈脂肪酸。脂肪酸根據碳氫鏈飽和與不飽和的不同可分為3類,即:飽和脂肪酸,碳氫上沒有不飽和鍵;單不飽和脂肪酸,其碳氫鏈有一個不飽和鍵;多不飽和脂肪酸,其碳氫鏈有二個或二個以上不飽和鍵。
脂肪酸是由碳、氫、氧三種元素組成的一類化合物,是中性脂肪、磷脂和糖脂的主要成分。
脂肪酸根據碳鏈長度的不同又可將其分為:
短鏈脂肪酸(short chain fatty acids,SCFA),其碳鏈上的碳原子數小於6,也稱作揮發性脂肪酸(volatile fatty acids,VFA);
中鏈脂肪酸(Midchain fatty acids,MCFA),指碳鏈上碳原子數為6-12的脂肪酸,主要成分是辛酸(C8)和癸酸(C10);
長鏈脂肪酸(Longchain fatty acids,LCFA),其碳鏈上碳原子數大於12。一般食物所含的大多是長鏈脂肪酸。
脂肪酸根據碳氫鏈飽和與不飽和的不同可分為3類,即:
飽和脂肪酸(Saturated fatty acids,SFA),碳氫上沒有不飽和鍵;
單不飽和脂肪酸(Monounsaturated fatty acids,MUFA),其碳氫鏈有一個不飽和鍵;
多不飽和脂肪酸(Polyunsaturated fatty acids,PUFA),其碳氫鏈有二個或二個以上不飽和鍵。
富含單不飽和脂肪酸和多不飽和脂肪酸組成的脂肪在室溫下呈液態,大多為植物油,如花生油、玉米油、豆油、堅果油(即阿甘油)、菜籽油等。以飽和脂肪酸為主組成的脂肪在室溫下呈固態,多為動物脂肪,如牛油、羊油、豬油等。但也有例外,如深海魚油雖然是動物脂肪,但它富含多不飽和脂肪酸,如20碳5烯酸(EPA)和22碳6烯酸(DHA),因而在室溫下呈液態。
CAS號:67254-79-9
脂肪酸可分成兩類:一類是分子內不帶碳碳雙鍵的飽和脂肪酸,如硬脂酸、軟脂酸等;另一類是分子內帶有一個或幾個碳碳雙鍵的不飽和脂肪酸,最常見的有油酸,油酸的碳鏈中只有一個碳碳雙鍵,所以又叫單不飽和脂肪酸。一般脂肪酸化合物的碳鏈都較短,其長度一般在18-36個碳原子,最少的就是12個碳原子,如月桂酸。不管飽和的或不飽和的,生物體內脂肪酸的碳原子數大多是偶數,極少含有奇數碳原子,尤其是在高等動植物體內主要存在12碳以上的高級脂肪酸,一般在14-24個碳,以16和18碳脂肪酸最為常見。奇數碳原子脂肪酸僅在一些植物、反芻動物、海洋生物、石油酵母等體內部分存在。
含有多量飽和脂肪酸的甘油i酯在常溫時往往是固體,例如牛油、羊油等,大多屬動物脂肪。含有較多不飽和脂肪酸的甘油三酯在常溫時往往是液體,例如玉米油、菜油等。植物和魚類的油大多是不飽和脂肪酸的甘油酯。
動物體內不能合成帶有2-4個雙鍵的不飽和脂肪酸,必須從食物中取得,因而這些脂肪酸就叫必需脂肪酸,也有人叫它維生素F。雖然已認為它們能降低血液中的膽固醇,但還沒有證據能證明人會因為食物中缺乏這些脂肪酸而引起疾病。微生物中也含有不飽和脂肪酸,藍細菌最獨特之處是含有兩個或多個雙鍵組成的不飽和脂肪酸,而細菌通常只含有飽和脂肪酸和一個雙鍵的不飽和脂肪酸。
脂肪酸(Fatty acid)具有長烴鏈的羧酸。通常以酯的形式為各種脂質的組分,以遊離形式存在的脂肪酸在自然界很罕見,最普通的脂肪酸見下表。
大多數脂肪酸含偶數碳原子,因為它們通常從2碳單位生物合成。高等動、植物最豐富的脂肪酸含16或18個碳原子,如棕櫚酸(軟脂酸)、油酸、亞油酸和硬脂酸。
動植物脂質的脂肪酸中超過半數為含雙鍵的不飽和脂肪酸,並且常是多雙鍵不飽和脂肪酸。細菌脂肪酸很少有雙鍵但常被羥化,或含有支鏈,或含有環丙烷的環狀結構。某些植物油和蠟含有不常見的脂肪酸。
不飽和脂肪酸必有1個雙鍵在C⑼和C⑽之間(從羧基碳原子數起)。脂肪酸的雙鍵幾乎總是順式幾何構型,這使不飽和脂肪酸的烴鏈有約30°的彎曲,干擾它們堆積時有效地填滿空間,結果降低了范德華相互反應力,使脂肪酸的熔點隨其不飽和度增加而降低。脂質的流動性隨其脂肪酸成分的不飽和度相應增加,這個現象對膜的性質有重要影響。
飽和脂肪酸是非常柔韌的分子,理論上圍繞每個C—C鍵都能相對自由地旋轉,因而有的構像範圍很廣。但是,其充分伸展的構象具有的能量最小,也最穩定;因為這種構象在毗鄰的亞甲基間的位阻最小。和大多數物質一樣,飽和脂肪酸的熔點隨分子重量的增加而增加。
動物能合成所需的飽和脂肪酸和油酸這類只含1個雙鍵的不飽和脂肪酸,含有2個或2個以上雙鍵的多雙鍵脂肪酸則必須從植物中獲取,故後者稱為必需脂肪酸,其中亞麻酸和亞油酸最重要。花生四烯酸從亞油酸生成。花生四烯酸是大多數前列腺素的前體,前列腺素是能調節細胞功能的激素樣物質。
自然界約有40多種不同的脂肪酸,它們是脂類的關鍵成分。許多脂類的物理特性取決於脂肪酸的飽和程度和碳鏈的長度,其中能為人體吸收、利用的只有偶數碳原子的脂肪酸。脂肪酸可按其結構不同進行分類,也可從營養學角度,按其對人體營養價值進行分類。按碳鏈長度不同分類。它可被分成短鏈(含2~4個碳原子)脂肪酸、中鏈(含6~12個碳原子)脂肪酸和長鏈(含14個以上碳原子)脂肪酸三類。人體內主要含有長鏈脂肪酸組成的脂類。
脂肪酸由C、H、O三種元素組成,是一端含有一個羧基的脂肪族碳氫鏈,是許多複雜酯的組成成分。低級脂肪酸是無色液體,有刺激性氣味,高級脂肪酸是蠟狀固體,無明顯氣味。脂肪酸在有充足氧供給的情況下,可氧化分解為CO2和H2O,釋放大量能量,因此是生物體的主要能量來源之一。
脂肪酸有多種分類形式,分別分類如下。
根據碳鏈長度的不同分類
可分為:短鏈脂肪酸、中鏈脂肪酸和長鏈脂肪酸。
脂肪酸代謝
脂肪酸根據碳鏈長度的不同又可將其分為:
短鏈脂肪酸(short chain fatty acids,SCFA),其碳鏈上的碳原子數小於6,也稱作揮發性脂肪酸(volatile fatty acids,VFA);
中鏈脂肪酸(Midchain fatty acids,MCFA),指碳鏈上碳原子數為6-12的脂肪酸,主要成分是辛酸(C8)和癸酸(C10);
長鏈脂肪酸(Longchain fatty acids,LCFA),其碳鏈上碳原子數大於12。一般食物所含的大多是長鏈脂肪酸。
根據碳氫鏈飽和與不飽和分類
脂肪酸根據碳氫鏈飽和與不飽和的不同可分為3類,即:
飽和脂肪酸(Saturated fatty acids,SFA),碳氫上沒有不飽和鍵;
單不飽和脂肪酸(Monounsaturated fatty acids,MUFA),其碳氫鏈有一個不飽和鍵;
多不飽和脂肪酸(Polyunsaturated fatty acids,PUFA),其碳氫鏈有二個或二個以上不飽和鍵。
(1)飽和脂肪酸碳氫鏈上沒有不飽和鍵,一般從C4到C38。從4個碳至24個碳原子的脂肪酸常存在於油脂中,而24個碳原子以上的則存在於蠟中。根據分子中碳原子數的多少可分為低級飽和脂肪酸(碳原子數≤10,常溫下為液態)和高級飽和脂肪酸(碳原子數>10,常溫下為固態)。動植物油脂中最常見的飽和脂肪酸有丁酸、己酸、辛酸、癸酸和高級飽和脂肪酸如十六酸(軟脂酸)與十八酸(硬脂酸),其次為十二酸(月桂酸)、十四酸(豆蔻酸)和二十酸(花生酸)等。
(2)不飽和脂肪酸分子中含有一個或一個以上不飽和鍵的脂肪酸都稱為不飽和脂肪酸。
不飽和脂肪酸通常呈液態,大多為植物油,如花生油、玉米油、豆油、堅果油(即阿甘油)、菜籽油等。根據不飽和鍵的多少又可分為單不飽和脂肪酸(有一個不飽和鍵,如豆蔻油酸、棕櫚油酸、菜籽油酸)和多不飽和脂肪酸(有兩個或兩個以上不飽和鍵,如亞油酸、亞麻酸)。不飽和脂肪酸以亞麻酸、亞油酸、油酸最為常見。
天然脂肪酸
現已發現一些多不飽和脂肪酸(從甲基端數起,最後一個不飽和雙鍵的位置在第三和第四個碳原子之間的脂肪酸)對人體有特殊的功能。最重要的這類脂肪酸是C22:6(4,7,10,13,16,19-二十二碳六烯酸,即DHA)和C20,5(5,8,11,14,17-二十碳五烯酸,即EPA),它們都屬於重要的功能性物質。研究表明:DHA有很好的健腦功能,並對老年性痴呆症、異位性皮炎、高脂血症有療效;EPA能使血小板凝聚能力降低、出血后血液凝固時間變長、心肌梗死發病率降低等。除上述功能外,EPA還可降低血液黏度、提高高密度膽固醇(優質膽固醇)的濃度,降低低密度膽固醇(劣質膽固醇)的濃度,因此EPA被認為可能對心血管疾病有良好的預防效果。DHA和EPA最主要的來源是深海魚油,如沙丁魚、烏賊、鱈魚等都有較多數量的DHA和EPA。
富含單不飽和脂肪酸和多不飽和脂肪酸組成的脂肪在室溫下呈液態,大多為植物油,如花生油、玉米油、豆油、堅果油(即阿甘油)、菜籽油等。以飽和脂肪酸為主組成的脂肪在室溫下呈固態,多為動物脂肪,如牛油、羊油、豬油等。但也有例外,如深海魚油雖然是動物脂肪,但它富含多不飽和脂肪酸,如EPA和DHA,因而在室溫下呈液態。
根據能夠滿足機體需要的程度分類
(1)必需脂肪酸
動物能合成所需的飽和脂肪酸和油酸這類只含1個雙鍵的不飽和脂肪酸,含有2個或2個以上雙鍵的多雙鍵脂肪酸則必須從植物中獲取,故後者稱為必需脂肪酸,其中亞麻酸和亞油酸最重要。花生四烯酸從亞油酸生成。花生四烯酸是大多數前列腺素的前體,前列腺素是能調節細胞功能的激素樣物質。
必需脂肪酸的最好來源是植物油,但在菜籽油和茶油中較其他植物油少。動物油脂中含量一般比植物油低,但相對來說,豬油比牛、羊脂含量多,禽類脂肪又比豬油多。肉類中雞、鴨肉較豬、牛、羊肉含量豐富。動物心、肝、腎和腸等內臟中的含量高於肌肉,而瘦肉中含量比肥肉多。此外,雞蛋黃中含量也較多。
(2)非必需脂肪酸
大多數脂肪酸人體能夠自身合成,可以不從食物中直接攝取,這類脂肪酸稱為非必需脂肪酸。非必需脂肪酸主要是飽和脂肪酸。雖然飽和脂肪酸為非必需脂肪酸,攝入過量會增加體內血脂的含量,但由於它對人體特別是對人的大腦的發育起著不可替代的作用,所以如果長期攝入不足,勢必會影響大腦的發育。因此應當根據實際情況來決定各種動物脂和植物油的攝入量。
它可分為飽和與不飽和脂肪酸兩大類。其中不飽和脂肪酸再按不飽和程度分為單不飽和脂肪酸與多不飽和脂肪酸。單不飽和脂肪酸,在分子結構中僅有一個雙鍵;多不飽和脂肪酸,在分子結構中含兩個或兩個以上雙鍵。
隨著營養科學的發展,發現雙鍵所在的位置影響脂肪酸的營養價值,因此又常按其雙鍵位置進行分類。雙鍵的位置可從脂肪酸分子結構的兩端第一個碳原子開始編號,並以其第一個雙鍵出現的位置的不同分別稱為ω-3族、ω-6族、ω-9族等不飽和脂肪酸。這一種分類方法在營養學上更有實用意義。
過去只重視ω-6族的亞油酸等,認為它們是必需脂肪酸,比較肯定的必需脂肪酸只有亞油酸。它們可由亞油酸轉變而成,在亞油酸供給充裕時這兩種脂肪酸即不至缺乏。自發現ω-3族脂肪酸以來,其生理功能及營養上的重要性越來越被人們重視。ω-3族脂肪酸包括麻酸及一些多不飽和脂肪酸,它們不少存在於深海魚的魚油中,其生理功能及營養作用有待開發與進一步研究。
必需脂肪酸不僅為營養所必需,而且與兒童生長發育和成長健康有關,更有降血脂、防治冠心病等治療作用,且與智力發育、記憶等生理功能有一定關係。
組成:
飽和脂肪酸(saturated fatty acid):不含有—C=C—雙鍵的脂肪酸。
不飽和脂肪酸(unsaturated fatty acid):至少含有—C=C—雙鍵的脂肪酸。
必需脂肪酸(essential fatty acid):維持哺乳動物正常生長所必需的,而動物又不能合成的脂肪酸,如亞油酸,亞麻酸。
三脂醯苷油(triacylglycerol):又稱為甘油三酯。一種含有與甘油脂化的三個脂醯基的酯。脂肪和油是三脂醯甘油的混合物。
磷脂(phospholipid):含有磷酸成分的脂。如卵磷脂,腦磷脂。
鞘脂(sphingolipid):一類含有鞘氨醇骨架的兩性脂,一端連接著一個長連的脂肪酸,另一端為一個極性和醇。鞘脂包括鞘磷脂,腦磷脂以及神經節苷脂,一般存在於植物和動物細胞膜內,猶其是在中樞神經系統的組織內含量豐富。
鞘磷脂(sphingomyelin):一種由神經醯胺的C-1羥基上連接了磷酸毛里求膽鹼(或磷酸乙醯胺)構成的鞘脂。鞘磷脂存在於在多數哺乳動物動物細胞的質膜內,是髓鞘的主要成分。
卵磷脂(lecithin):即磷脂醯膽鹼(PC),是磷脂醯與膽鹼形成的複合物。
腦磷脂(cephalin):即磷脂醯乙醇胺(PE),是磷脂醯與乙醇胺形成的複合物。
脂質體(liposome):是由包圍水相空間的磷脂雙層形成的囊泡(小泡)。
人體各組織脂肪中脂肪酸的含量
常用食用油脂中主要脂肪酸的組成
酮體(acetone bodies)是脂肪酸在肝臟進行正常分解代謝所生成的特殊中間產物,包括有乙醯乙酸(acetoacetic acid約佔30%),β-羥丁酸(β?hydroxybutyric acid約佔70%)和極少量的丙酮(acetone)(分子式見下圖)。正常人血液中酮體含量極少,這是人體利用脂肪氧化供能的正常現象。但在某些生理情況(飢餓、禁食)或病理情況下(如糖尿病),糖的來源或氧化供能障礙,脂動員增強,脂肪酸就成了人體的主要供能物質。若肝中合成酮體的量超過肝外組織利用酮體的能力,二者之間失去平衡,血中濃度就會過高,導致酮血症(acetonemia)和酮尿症(acetonuria)。乙醯乙酸和β-羥丁酸都是酸性物質,因此酮體在體內大量堆積還會引起酸中毒。
⒈乙醯CoA的轉移
乙醯CoA可由糖氧化分解或由脂肪酸、酮體和蛋白分解生成,生成乙醯CoA的反應均發生在線粒體中,而脂肪酸的合成部位是胞漿,因此乙醯CoA必須由線粒體轉運至胞漿。但是乙醯CoA不能自由通過線粒體膜,需要通過一個稱為檸檬酸—丙酮酸循環(citrate pyruvate cycle)來完成乙醯CoA由線粒體到胞漿的轉移。
首先在線粒體內,乙醯CoA與草醯乙酸經檸檬酸合成酶催化,縮合生成檸檬酸,再由線粒體內膜上相應載體協助進入胞液,在胞液內存在的檸檬酸裂解酶(citrate lyase)可使檸檬酸裂解產生乙醯CoA及草醯乙酸。前者即可用於生成脂肪酸,後者可返回線粒體補充合成檸檬酸時的消耗。但草醯乙酸也不能自由通透線粒體內膜,故必須先經蘋果酸脫氫酶催化,還原成蘋果酸再經線粒體內膜上的載體轉運入線粒體,經氧化後補充草醯乙酸。也可在蘋果酸酶作用下,氧化脫羧生成丙酮酸,同時伴有NADPH的生成。丙酮酸可經內膜載體被轉運入線粒體內,此時丙酮酸可再羧化轉變為草醯乙酸。每經檸檬酸丙酮酸循環一次,可使一分子乙酸CoA由線粒體進入胞液,同時消耗兩分子ATP,還為機體提供了NADPH以補充合成反應的需要。
⒉丙二醯CoA的生成
乙醯CoA由乙醯CoA羧化酶(acetyl CoA carboxylase)催化轉變成丙二醯CoA(或稱丙二酸單醯CoA),乙醯CoA羧化酶存在於胞液中,其輔基為生物素,在反應過程中起到攜帶和轉移羧基的作用。該反應機理類似於其他依賴生物素的羧化反應,如催化丙酮酸羧化成為草醯乙酸的反應等。反應如下:
由乙醯CoA羧化酶催化的反應為脂肪酸合成過程中的限速步驟。此酶為一別構酶,在變構效應劑的作用下,其無活性的單體與有活性的多聚體(由100個單體呈線狀排列)之間可以互變。檸檬酸與異檸檬酸可促進單體聚合成多聚體,增強酶活性,而長鏈脂肪酸可加速解聚,從而抑制該酶活性。乙醯CoA羧化酶還可通過依賴於cAMP的磷酸化及去磷酸化修飾來調節酶活性。此酶經磷酸化后活性喪失,如胰高血糖素及腎上腺素等能促進這種磷酸化作用,從而抑制脂肪酸合成;而胰島素則能促進酶的去磷酸化作用,故可增強乙醯CoA羧化酶活性,加速脂肪酸合成。
同時乙醯CoA羧化酶也是誘導酶,長期高糖低脂飲食能誘導此酶生成,促進脂肪酸合成;反之,高脂低糖飲食能抑制此酶合成,降低脂肪酸的生成。
⒊軟脂酸的生成
在原核生物(如大腸桿菌中)催化脂肪酸生成的酶是一個由7種不同功能的酶與一種醯基載體蛋白(acyl carrier protein,ACP)聚合成的複合體。在真核生物催化此反應是一種含有雙亞基的酶,每個亞基有7個不同催化功能的結構區和一個相當於ACP的結構區,因此這是一種具有多種功能的酶。不同的生物此酶的結構有差異。
軟脂酸的合成實際上是一個重複循環的過程,由1分子乙醯CoA與7分子丙二醯CoA經轉移、縮合、加氫、脫水和再加氫重複過程,每一次使碳鏈延長兩個碳,共7次重複,最終生成含十六碳的軟脂酸。
脂肪酸合成需消耗ATP和NADPH+H+,NADPH主要來源於葡萄糖分解的磷酸戊糖途徑。此外,蘋果酸氧化脫羧也可產生少量NADPH。
脂肪酸合成過程不是β-氧化的逆過程,它們反應的組織,細胞定位,轉移載體,醯基載體,限速酶,激活劑,抑製劑,供氫體和受氫體以及反應底物與產物均不相同。
機體內不僅有軟脂酸,還有碳鏈長短不等的其它脂肪酸,也有各種不飽和脂肪酸,除營養必需脂肪酸依賴食物供應外,其它脂肪酸均可由軟脂酸在細胞內加工改造而成。
⒈碳鏈的延長和縮短
脂肪酸碳鏈的縮短在線粒體中經β-氧化完成,經過一次β-氧化循環就可以減少兩個碳原子。
脂肪酸碳鏈的延長可在滑面內質網和線粒體中經脂肪酸延長酶體系催化完成。
在內質網,軟脂酸延長是以丙二醯CoA為二碳單位的供體,由NADPH+H+供氫,亦經縮合脫羧、還原等過程延長碳鏈,與胞液中脂肪酸合成過程基本相同。但催化反應的酶體系不同,其脂肪醯基不是以ACP為載體,而是與輔酶A相連參加反應。除腦組織外一般以合成硬脂酸(18C)為主,腦組織因含其他酶,故可延長至24碳的脂肪酸,供腦中脂類代謝需要。
在線粒體,軟脂酸經線粒體脂肪酸延長酶體系作用,與乙醯CoA縮合逐步延長碳鏈,其過程與脂肪酸β氧化逆行反應相似,僅烯脂醯CoA還原酶的輔酶為NADPH+H+與β氧化過程不同。通過此種方式一般可延長脂肪酸碳鏈至24或26碳,但以硬脂酸最多。
⒉脂肪酸脫飽和
人和動物組織含有的不飽和脂肪酸主要為軟油酸(16:1△9)、油酸(18:1△9)、亞油酸(18:2△9,12)、亞麻酸(18:3△9,12,15)、花生四烯酸(20:4△5,8,11,14)等。其中最普通的單不飽和脂肪酸軟油酸和油酸可由相應的脂肪酸活化后經去飽和酶(acylCoAdesaturase)催化脫氫生成。這類酶存在於滑面內質網,屬混合功能氧化酶;因該酶只催化在△9形成雙鍵,而不能在C10與末端甲基之間形成雙鍵,故亞油酸(linoleate)、亞麻酸(linolenate)及花生四烯酸(arachidonate)在體內不能合成或合成不足。但它們又是機體不可缺少的,所以必須由食物供給,因此,稱之為必需脂肪酸(essential fatty acid)。
植物組織含有可以在C-10與末端甲基間形成雙鍵(即ω3和ω6)的去飽和酶,能合成以上3種多不飽和脂肪酸。當食入亞油酸后,在動物體內經碳鏈加長及去飽和后,可生成花生四烯酸。
乙醯CoA羧化酶催化的反應是脂肪酸合成的限速步驟,很多因素都可影響此酶活性,從而使脂肪酸合成速度改變。脂肪酸合成過程中其他酶,如脂肪酸合成酶、檸檬酸裂解酶等亦可被調節。
⒈代謝物的調節
在高脂膳食后,或因飢餓導致脂肪動員加強時,細胞內軟脂醯CoA增多,可反饋抑制乙醯CoA羧化酶,從而抑制體內脂肪酸合成。而進食糖類,糖代謝加強時,由糖氧化及磷酸戊糖循環提供的乙醯CoA及NADPH增多,這些合成脂肪酸的原料的增多有利於脂肪酸的合成。此外,糖氧化加強的結果,使細胞內ATP增多,進而抑制異檸檬酸脫氫酶,造成異檸檬酸及檸檬酸堆積,在線粒體內膜的相應載體協助下,由線粒體轉入胞液,可以別構激活乙醯CoA羧化酶。同時本身也可裂解釋放乙醯CoA,增加脂肪酸合成的原料,使脂肪酸合成增加。
⒉激素的調節
胰島素、胰高血糖素、腎上腺素及生長素等均參與對脂肪酸合成的調節。
胰島素能誘導乙醯CoA羧化酶、脂肪酸合成酶及檸檬酸裂解酶的合成,從而促進脂肪酸的合成。此外,還可通過促進乙醯CoA羧化酶的去磷酸化而使酶活性增強,也使脂肪酸合成加速。
胰高血糖素等可通過增加cAMP,致使乙醯CoA羧化酶磷酸化而降低活性,因此抑制脂肪酸的合成。此外,胰高血糖素也抑制甘油三酯合成,從而增加長鏈脂醯CoA對乙醯CoA羧化酶的反饋抑制,亦使脂肪酸合成被抑制。
無論是植物性或動物性油脂每克都有9卡的熱量。但是植物性油含分解脂肪的物質,適度攝取是有益的,但並不表示其熱量較低。一般人認為植物油很安全,可以多吃,這個是錯誤的觀念,不但減肥的人必須限量攝食植物油,以免對減肥不利,要健康長壽的人更應如此。
人們所需的脂肪酸有三類:多元不飽和脂肪酸、單元不飽和脂肪酸和飽和脂肪酸。我們常用的食用油通常都含人體需要的三種脂肪酸。
每人每日油脂攝取量只能占每日食物總熱量的二成,(每天的用油量控制在15至30毫升)每人每天要吃齊這三種脂肪酸,不能偏好任一油類,否則油脂攝取失衡,會形成疾病。每日單元不飽和脂肪酸的攝食量要佔一成,多元不飽和脂肪酸要佔一成,而飽和脂肪酸要少於一成。
動物油、椰子油和棕櫚油的主要成分是飽和脂肪酸,而多元不飽和脂肪酸的含量很低。心臟病人捨棄動物性飽和油后,可從植物油中攝取植物性飽和油。
橄欖油、堅果油(即阿甘油)、菜籽油、玉米油、花生油的單元不飽和脂肪酸含量較高,人體需要的三種脂肪酸中,以單元不飽和脂肪酸的需要量最大,玉米油、橄欖油可作這種脂肪酸的重要來源。
葵花油、粟米油油、大豆等植物油和海洋魚類中含的脂肪多為多元不飽和脂肪酸。多元不飽和脂肪酸是這些食用油的主要成份,其他兩種脂肪酸含量不多。三種脂肪酸中,多元不飽和脂肪酸最不穩定,在油炸、油炒或油煎的高溫下,最容易被氧化變成毒油。而偏偏多元不飽和脂肪酸又是人體細胞膜的重要原料之一。在細胞膜內也有機會被氧化,被氧化后,細胞膜會喪失正常機能而使人生病。故即使不吃動物油而只吃植物油,吃得過量,也一樣會增加得大腸乳癌、直腸癌、前列腺癌或其他疾病的機會。
高油脂食物是人們得癌症的重要成因之一,而癌症又是人類死亡的主要原因之一,隨著人們物質的富裕,大家的脂肪攝入量也正在逐年增加,預期在往後幾十年裡,人們得癌症的可能性也將逐年增加。癌症的形成需要十五至四十五年,過程非常緩慢,以前癌症發生都在中老年人身上,已有年輕化的跡象,所以我們要養成少吃油脂的習慣,讓自己現在苗條,健康。
當然,在現代人們生活條件不斷提升,脂肪酸的攝入量由無法控制的情況下,可定期食用魔芋膳食纖維,平衡人體營養所需。從而改善人體腸道、並分解和排除過量的脂肪酸,增加人們的長壽和控制癌症的發病率。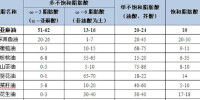
脂肪酸
來源
人們對脂肪酸的研究中發現,有的脂肪酸分子結構中含有“雙鍵”,有的不含雙鍵,人們把含雙鍵的脂肪酸叫不飽和脂肪酸,把不含雙鍵的叫飽和脂肪酸。大多數植物油含不飽和脂肪酸較多,如大豆油、花生油、芝麻油、玉米油、阿甘油、葵花子油含量較多,而動物油含不飽和脂肪酸很低。奶油含有的不飽和脂肪酸亦低,但含有維生素A、D,溶點低,易於消化,小兒可以食用。脂肪中所含不飽和脂肪酸有油酸、亞油酸、亞麻油酸、花生四烯酸等。但有的不飽和脂肪人體可以合成,有不能合成。
脂肪酸
各類碳鏈長短脂肪酸名稱:
● C6酸己酸
● C8酸辛酸
● C10酸癸酸
● C12酸月桂酸
● C14酸肉豆蔻酸
● C16酸棕櫚酸
● C18酸硬脂酸
● C20酸花生酸
● C22酸山嵛酸
● C24酸木質素酸
● C26酸蠟酸
● C28酸褐煤酸
● C30酸蜜蠟酸
(1)色澤與氣味
純凈的脂肪酸是無色的,某些脂肪酸具有自己特有的氣味。
(2)密度
脂肪酸的相對密度一般都小於1,與其相對分子質量成反比,隨溫度的升高而降低,隨碳鏈增長而減小,不飽和鍵越多密度越大。
(3)熔點
脂肪酸的熔點隨著碳鏈的增長呈不規則升高,奇數碳原子鏈脂肪酸的熔點低於其相鄰的偶數碳脂肪酸,不飽和脂肪酸的熔點通常低於飽和脂肪酸,雙鍵越多,熔點越低,雙鍵位置越靠近碳鏈兩端,熔點越高。
引入一個雙鍵到碳鏈中會降低脂肪酸的熔點,雙鍵位置越向碳鏈中部移動,熔點降低越大,順式雙鍵產生的這種影響大於反式。雙鍵增加熔點下降,但共軛雙鍵不在此例。經過氫化、反化或非共軛雙鍵異構化成共軛烯酸等都會提高熔點。每一個奇數碳原子脂肪酸的熔點,小於與它最接近的偶數碳原子脂肪酸的熔點,例如十七酸的熔點( 61.3℃),既低於十八酸的(69.6℃),也低於十六酸的(62.7℃)。此現象不僅存在於脂肪酸,也見於其他長碳鏈化合物。
(4)沸點
脂肪酸的沸點隨碳鏈增長而升高,飽和度不同但碳鏈長度相同的脂肪酸沸點相近。
(5)溶解性
低級脂肪酸易溶於水,但隨著相對分子質量的增加,在水中的溶解度減小,以至溶或不溶於水,而溶於有機溶劑。一般脂肪酸越低級,不飽和度越高,其在有機溶劑中的溶解度也就越大,溫度越高溶解度越大,碳鏈越長溶解度越小。
物質的物理性質,是其化學組成與結構的表現。在高級脂肪酸中,存在非極性的長碳鏈和極性的-COOH基與-COOR基。碳鏈長短與不飽和鍵的多少各有差異,導致脂肪酸的各種物理與化學性質的差異有的很小,有的很大,有時微小的差別顯示出重大的意義。
脂肪酸可以用下面的方式表達它們的名稱、碳原子數、不飽和雙鍵的數目和位置。在表達它們的名稱時,先寫出碳原子的數目,再寫出雙鍵的數目,最後用△及右上角的數字錶示雙鍵的位置,並在雙鍵位置數字後面加上c(cis,順式)或t(trans,反式)表示雙鍵的構型。例如,亞油酸的化學名稱是順,順-9,12-十八烯酸。
天然脂肪酸的分子結構存在一些共同規律:
(1)一般都是碳數為偶數的長鏈脂肪酸,14-20個碳原子的佔多數,最常見的是16或18個碳原子數的,如軟脂酸(16:0)、硬脂酸(18:0)和油酸(18:1△)。
(2)高等動植物的不飽和脂肪酸一般都是順式結構(cis),反式(trans)很少。
(3)不飽和脂肪酸的雙鍵位置有一定的規律:一個雙鍵者,位置在9和10碳原子之間,多個雙鍵者,也常有9位的雙鍵,其餘雙鍵在C。與碳鏈甲基末端之間,兩個雙鍵之間有亞甲基間隔,如油酸(18:1△)、亞油酸(18:2△)、亞麻酸(18:3△)、花生四烯酸(20:4△)。
(4)一般動物脂肪中含飽和脂肪酸多;而高等植物和在低溫條件下生長的動物的脂肪中,不飽和脂肪酸的含量較高。
天然三醯基甘油的飽和脂肪酸絕大多數都是偶碳數直鏈的,奇碳數鏈的極個別,含量也極少。
飽和脂肪酸是非常柔韌的分子,理論上圍繞每個C-C鍵都能相對自由地旋轉,因而有的構像範圍很廣。但是,其充分伸展的構象具有的能量最小,也最穩定;因為這種構象在毗鄰的亞甲基間的位阻最小。和大多數物質一樣,飽和脂肪酸的熔點隨分子重量的增加而增加。
動植物脂質的脂肪酸中超過半數為含雙鍵的不飽和脂肪酸,並且常是多雙鍵不飽和脂肪酸。細菌脂肪酸很少有雙鍵但常被羥化,或含有支鏈,或含有環丙烷的環狀結構。某些植物油和蠟含有不常見的脂肪酸。
哺乳動物和人體不能合成亞油酸和亞麻酸,而它們又是生長所必需的,需要由食物供給,故稱為必需脂肪酸( essential fatty acids)。這兩種脂肪酸在植物中含量非常豐富,哺乳動物中的花生四烯酸是由亞油酸合成的,花生四烯酸在植物中含量很少。
氧化分解
β-氧化
脂肪酸不溶於水,在血液中與清蛋白結合后(10:1),運送至全身各組織細胞,在細胞的線粒體內氧化分解,釋放出大量能量,以肝臟和肌肉最為活躍。1904年,Knoop剛苯環作標記,追蹤脂肪酸在動物體內的轉變,發現奇數碳脂肪酸衍生物被降解時,尿中檢出馬尿酸,若是偶數碳,尿中檢出苯乙尿酸。推測脂肪酸醯基鏈的降解發生在β-碳原子上,即每次從脂酸鏈上切下一個二碳單位。後來的實驗證明β-氧化學說是正確的,切下的二碳單位是乙醯CoA,脂肪酸進入線粒體前要先被活化。
β-氧化
1)脂肪酸的活化;
2)脂醯CoA進入線粒體;
3)脂醯CoA的β-氧化;
脂醯CoA氧化生成乙醯CoA涉及四個反應—脫氫、加水、再脫氫、硫解。每一次產生1分子乙醯CoA和比原來少2個C的脂醯CoA。再進行下一輪β-氧化,如此循環反覆。
4)脂肪酸氧化的能量計算
以脂肪為能源時,生物體還可獲得大量的水。駱駝的駝峰是儲存脂的“倉庫”,既可提供能量,又可提供所需的水。
脂肪酸氧化的其他途徑
(1)奇數碳原子脂肪酸的氧化。人體含微量奇數碳脂肪酸,許多植物、海洋生物和石油酵母等含一定量的奇數碳脂肪酸。其β-氧化除生成乙醯CoA外,還生成1分子丙醯CoA,後者在β-羧化酶及異構酶的作用下生成琥珀醯CoA,經TCA途徑徹底氧化。
(2)不飽和脂肪酸的氧化。機體中約一半以上的脂肪酸是不飽和脂肪酸,其中雙鍵均為順式(cis)構型,它不能被烯脂醯CoA水化酶催化,該酶催化反式構型雙鍵的加水過程,所以需要異構酶和還原酶參與才能使一般不飽和脂肪酸的氧化進行下去。如油酸是十八碳一烯酸((cis-△),細胞質中的油酸同樣先活化生成油醯CoA,經轉運系統生成線粒體基質中的油醯CoA,經三輪β-氧化生成3分子乙醯CoA和cis-△-十二碳烯脂醯CoA,後者經異構酶轉化為trans-△一十二碳烯脂醯CoA,由烯脂醯CoA水化酶作用生成L-β-羥脂醯CoA,再經五輪β-氧化生成6分子乙醯CoA,總計生成9分子乙醯CoA。
多不飽和脂肪酸的氧化還需一個特殊的還原酶參與進行。
酮體
酮體(acetone bodies)是脂肪酸在肝臟進行正常分解代謝所生成的特殊中間產物,包括有乙醯乙酸(acetoacetic acid約佔30%),β-羥丁酸(β?hydroxybutyric acid約佔70%)和極少量的丙酮(acetone)。正常人血液中酮體含量極少,這是人體利用脂肪氧化供能的正常現象。但在某些生理情況(飢餓、禁食)或病理情況下(如糖尿病),糖的來源或氧化供能障礙,脂動員增強,脂肪酸就成了人體的主要供能物質。若肝中合成酮體的量超過肝外組織利用酮體的能力,二者之間失去平衡,血中濃度就會過高,導致酮血症(acetonemia)和酮尿症(acetonuria)。乙醯乙酸和β-羥丁酸都是酸性物質,因此酮體在體內大量堆積還會引起酸中毒。
生物合成
脂肪酸合成部位
體內肝、腎、腦、肺、乳腺、脂肪等組織的細胞質中均存在脂肪酸的合成酶系,因此這些組織均能合成脂肪酸,但以肝的脂肪酸合成酶系活性最高,閃此肝細胞是人體內合成脂肪酸的主要部位。
脂肪組織雖然也能以葡萄糖代謝的中間產物為原料合成脂肪酸,其主要來源是小腸吸收的外源性脂肪酸和肝合成的內源性脂肪酸。
脂肪酸合成
軟脂酸的合成是在細胞質完成,但脂肪酸鏈延長則是在線粒體和內質網完成。
脂肪酸合成原料
合成脂肪酸的原料有乙醯輔酶A、HCO3(C02)、NADPH和ATP,Mn可作為酶的激活劑。
脂肪酸合成途徑
生物體內由乙醯CoA合成脂肪酸的有:①非線粒體酶系合成途徑:即胞漿酶系合成飽和脂肪酸途徑。該途徑的終產物是軟脂酸,故又稱為軟脂酸合成途徑,它是脂肪酸合成的主要途徑。②線粒體酶系合成途徑:又稱飽和脂肪酸碳鏈延長途徑。
軟脂酸
1.乙醯CoA的轉移
乙醯CoA可由糖氧化分解或由脂肪酸、酮體和蛋白分解生成,生成乙醯CoA的反應均發生在線粒體中,而脂肪酸的合成部位是胞漿,因此乙醯CoA必須由線粒體轉運至胞漿。但是乙醯CoA不能自由通過線粒體膜,需要通過一個稱為檸檬酸-丙酮酸循環(citrate pyruvate cycle)來完成乙醯CoA由線粒體到胞漿的轉移。
首先在線粒體內,乙醯CoA與草醯乙酸經檸檬酸合成酶催化,縮合生成檸檬酸,再由線粒體內膜上相應載體協助進入胞液,在胞液內存在的檸檬酸裂解酶(citrate lyase)可使檸檬酸裂解產生乙醯CoA及草醯乙酸。前者即可用於生成脂肪酸,後者可返回線粒體補充合成檸檬酸時的消耗。但草醯乙酸也不能自由通透線粒體內膜,故必須先經蘋果酸脫氫酶催化,還原成蘋果酸再經線粒體內膜上的載體轉運入線粒體,經氧化後補充草醯乙酸。也可在蘋果酸酶作用下,氧化脫羧生成丙酮酸,同時伴有NADPH的生成。丙酮酸可經內膜載體被轉運入線粒體內,此時丙酮酸可再羧化轉變為草醯乙酸。每經檸檬酸丙酮酸循環一次,可使一分子乙酸CoA由線粒體進入胞液,同時消耗兩分子ATP,還為機體提供了NADPH以補充合成反應的需要。
2.丙二醯CoA的生成
乙醯CoA由乙醯CoA羧化酶(acetyl CoA carboxylase)催化轉變成丙二醯CoA(或稱丙二酸單醯CoA),乙醯CoA羧化酶存在於胞液中,其輔基為生物素,在反應過程中起到攜帶和轉移羧基的作用。該反應機理類似於其他依賴生物素的羧化反應,如催化丙酮酸羧化成為草醯乙酸的反應等。
由乙醯CoA羧化酶催化的反應為脂肪酸合成過程中的限速步驟。此酶為一別構酶,在變構效應劑的作用下,其無活性的單體與有活性的多聚體(由100個單體呈線狀排列)之間可以互變。檸檬酸與異檸檬酸可促進單體聚合成多聚體,增強酶活性,而長鏈脂肪酸可加速解聚,從而抑制該酶活性。乙醯CoA羧化酶還可通過依賴於cAMP的磷酸化及去磷酸化修飾來調節酶活性。此酶經磷酸化后活性喪失,如胰高血糖素及腎上腺素等能促進這種磷酸化作用,從而抑制脂肪酸合成;而胰島素則能促進酶的去磷酸化作用,故可增強乙醯CoA羧化酶活性,加速脂肪酸合成。
同時乙醯CoA羧化酶也是誘導酶,長期高糖低脂飲食能誘導此酶生成,促進脂肪酸合成;反之,高脂低糖飲食能抑制此酶合成,降低脂肪酸的生成。
3.軟脂酸的生成
脂肪酸分離設備
在原核生物(如大腸桿菌中)催化脂肪酸生成的酶是一個由7種不同功能的酶與一種醯基載體蛋白(acyl carrier protein,ACP)聚合成的複合體。在真核生物催化此反應是一種含有雙亞基的酶,每個亞基有7個不同催化功能的結構區和一個相當於ACP的結構區,因此這是一種具有多種功能的酶。不同的生物此酶的結構有差異。
軟脂酸的合成實際上是一個重複循環的過程,由1分子乙醯CoA與7分子丙二醯CoA經轉移、縮合、加氫、脫水和再加氫重複過程,每一次使碳鏈延長兩個碳,共7次重複,最終生成含十六碳的軟脂酸。
脂肪酸合成需消耗ATP和NADPH+H+,NADPH主要來源於葡萄糖分解的磷酸戊糖途徑。此外,蘋果酸氧化脫羧也可產生少量NADPH。
脂肪酸合成過程不是β-氧化的逆過程,它們反應的組織,細胞定位,轉移載體,醯基載體,限速酶,激活劑,抑製劑,供氫體和受氫體以及反應底物與產物均不相同。
其它脂酸類
機體內不僅有軟脂酸,還有碳鏈長短不等的其它脂肪酸,也有各種不飽和脂肪酸,除營養必需脂肪酸依賴食物供應外,其它脂肪酸均可由軟脂酸在細胞內加工改造而成。
1.碳鏈的延長和縮短
脂肪酸碳鏈的縮短在線粒體中經β-氧化完成,經過一次β-氧化循環就可以減少兩個碳原子。
脂肪酸碳鏈的延長可在滑面內質網和線粒體中經脂肪酸延長酶體系催化完成。
在內質網,軟脂酸延長是以丙二醯CoA為二碳單位的供體,由NADPH+H+供氫,亦經縮合脫羧、還原等過程延長碳鏈,與胞液中脂肪酸合成過程基本相同。但催化反應的酶體系不同,其脂肪醯基不是以ACP為載體,而是與輔酶A相連參加反應。除腦組織外一般以合成硬脂酸(18C)為主,腦組織因含其他酶,故可延長至24碳的脂肪酸,供腦中脂類代謝需要。
在線粒體,軟脂酸經線粒體脂肪酸延長酶體系作用,與乙醯CoA縮合逐步延長碳鏈,其過程與脂肪酸β氧化逆行反應相似,僅烯脂醯CoA還原酶的輔酶為NADPH+H+與β氧化過程不同。通過此種方式一般可延長脂肪酸碳鏈至24或26碳,但以硬脂酸最多。
2.脂肪酸脫飽和
人和動物組織含有的不飽和脂肪酸主要為軟油酸(16:1△9)、油酸(18:1△9)、亞油酸(18:2△9,12)、亞麻酸(18:3△9,12,15)、花生四烯酸(20:4△5,8,11,14)等。其中最普通的單不飽和脂肪酸軟油酸和油酸可由相應的脂肪酸活化后經去飽和酶(acylCoAdesaturase)催化脫氫生成。這類酶存在於滑面內質網,屬混合功能氧化酶;因該酶只催化在△9形成雙鍵,而不能在C10與末端甲基之間形成雙鍵,故亞油酸(linoleate)、亞麻酸(linolenate)及花生四烯酸(arachidonate)在體內不能合成或合成不足。但它們又是機體不可缺少的,所以必須由食物供給,因此,稱之為必需脂肪酸(essential fatty acid)。
植物組織含有可以在C-10與末端甲基間形成雙鍵(即ω3和ω6)的去飽和酶,能合成以上3種多不飽和脂肪酸。當食入亞油酸后,在動物體內經碳鏈加長及去飽和后,可生成花生四烯酸。
調節方法
乙醯CoA羧化酶催化的反應是脂肪酸合成的限速步驟,很多因素都可影響此酶活性,從而使脂肪酸合成速度改變。脂肪酸合成過程中其他酶,如脂肪酸合成酶、檸檬酸裂解酶等亦可被調節。
1.代謝物的調節
在高脂膳食后,或因飢餓導致脂肪動員加強時,細胞內軟脂醯CoA增多,可反饋抑制乙醯CoA羧化酶,從而抑制體內脂肪酸合成。而進食糖類,糖代謝加強時,由糖氧化及磷酸戊糖循環提供的乙醯CoA及NADPH增多,這些合成脂肪酸的原料的增多有利於脂肪酸的合成。此外,糖氧化加強的結果,使細胞內ATP增多,進而抑制異檸檬酸脫氫酶,造成異檸檬酸及檸檬酸堆積,在線粒體內膜的相應載體協助下,由線粒體轉入胞液,可以別構激活乙醯CoA羧化酶。同時本身也可裂解釋放乙醯CoA,增加脂肪酸合成的原料,使脂肪酸合成增加。
2.激素的調節
胰島素、胰高血糖素、腎上腺素及生長素等均參與對脂肪酸合成的調節。
胰島素能誘導乙醯CoA羧化酶、脂肪酸合成酶及檸檬酸裂解酶的合成,從而促進脂肪酸的合成。此外,還可通過促進乙醯CoA羧化酶的去磷酸化而使酶活性增強,也使脂肪酸合成加速。
胰高血糖素等可通過增加cAMP,致使乙醯CoA羧化酶磷酸化而降低活性,因此抑制脂肪酸的合成。此外,胰高血糖素也抑制甘油三酯合成,從而增加長鏈脂醯CoA對乙醯CoA羧化酶的反饋抑制,亦使脂肪酸合成被抑制。
脂肪酸常與其他物質結合形成酯,以遊離形式存在的脂肪酸在自然界很罕見。
人在遇到飢餓或壓力時,激素會激活脂肪細胞中的脂肪酶,將儲存的甘油三酯轉變回脂肪酸和甘油,然行它們被釋放到血液中得到利用。除了腦細胞之外,身體的所有細胞在飢餓缺乏能量剛‘都使自己適應於利用脂肪酸,脂肪酸同葡萄糖一樣可轉化成ATP的能量形式。事實上,能刺激甘油三酯裂解的激素在大腦內卻是無效的。人的大腦由於不具備像身體其他部位那樣的利用脂肪酸的能力,它只有利用葡萄糖。甘油三酯裂解后的另一產物-甘油,則循環至肝臟,肝臟將其通過另一條生物化學途徑轉化成葡萄糖以供養大腦。就這樣,當養料缺乏時,身體的其他部分可依賴於脂肪酸,而大腦只能依靠它所需要得到的葡萄糖。
天然脂肪酸
動物能合成所需的飽和脂肪酸和亞油酸這類只含1個雙鍵的不飽和脂肪酸,含有2個或2個以上雙鍵的多雙鍵脂肪酸則必須從植物中獲取,稱為必需脂肪酸,其中亞麻酸和亞油酸最重要。在大部分含油脂豐富的食物中,有一半左右的熱量是由脂肪和油類提供的。天然的脂肪和油類通常是由一種以上的脂肪酸與甘油形成的各種酯的混合物。這些脂肪酸的功能如下。
①能提供熱量,是很好的能量來源。
②脂肪酸貯存在脂肪細胞中,以備人體不時之需。
③作為合成其他化合物的原料。
④能保持細胞膜的相對流動性,以保證細胞的正常生理功能。
⑤使膽固醇酯化,降低血液中膽固醇和甘油三酯含量。
⑥提高腦細胞活性,增強記憶力和思維能力。
脂肪酸可用於丁苯橡膠生產中的乳化劑和其它表面活性劑、潤滑劑、光澤劑;還可用於生產高級香皂、透明皂、硬脂酸及各種表面活性劑的中間體。
每一類、每一種脂肪酸均有其特定用途和功能特性。功能性脂肪酸是特指那些來源於人類膳食油脂,為人體營養、健康所必需,並對現已發現的人體一些相應缺乏症和內源性疾病,特別是對現今社會文明病如高血壓、心臟病、癌症、糖尿病等有積極防治作用的一組脂肪酸,這其中又以備受關注和廣為研究的多不飽和脂肪酸為主。
多不飽和脂肪酸
多不飽和脂肪酸是功能性脂肪酸研究和開發的主體與核心,根據其結構又分為n-6和n-3兩大主要系列。這類脂肪酸受到廣泛關注,不僅僅因為n-6系列的亞油酸和n-3系列的α-亞麻酸是人體不可或缺的必需脂肪酸,更重要的是因為其在人體生理中起著極為重要的代謝作用,與現代諸多文明病的發生與調控息息相關。目前認為n-6和n-3脂肪酸功能的突出重要性,首先在於它們是體內有重要代謝功能的前列腺素、白三烯等的前體。另一突出重要性在於,它們是人體器官和組織生物膜的絕對必需成分。此外,這些多不飽和脂肪酸分子本身在人體其他許多正常生理過程中起著特殊作用。
1.亞油酸
亞油酸(linoleic acid)是功能性多不飽和脂肪酸中被最早認識的一種,而且在世界範圍內的絕大多數膳食營養中佔據著不飽和脂肪酸的大部分。亞油酸具有降低血清膽固醇水平的作用,與12:0-16:0飽和脂肪酸相比,亞油酸具有較強的降低LDL-膽固醇的濃度的作用。攝入大量亞油酸對高三醯基甘油血症病人效果較為明顯。我國藥典仍有採用亞油酸乙酯丸劑、滴劑作預防和治療高血壓及動脈粥樣硬化症、冠心病的藥物。
2.花生四烯酸
亞油酸被定為必需脂肪酸的部分原因在於它是n-6長鏈多不飽和脂肪酸,還是花生四烯酸(arachidonic acid)的前體,花生四烯酸較多地存在於神經組織和腦中,大腦積極地代謝花生四烯酸,其代謝產物對中樞神經系統有重要影響,包括神經元跨膜信號的調整、神經遞質的釋放以及葡萄糖的攝取。從妊娠的第三個月到約2歲嬰兒的生命成長發育中,花生四烯酸在大腦內快速積累,在細胞分裂和信號傳遞方面起重要作用。對於成年人,膳食花生四烯酸的供給是否影響與腦代謝有關的花生四烯酸底物庫尚不清楚。在一些抗腫瘤動物試驗中,已證明花生四烯酸在體外能顯著殺滅腫瘤細胞,而且對正常細胞沒有顯示出毒副作用。花生四烯酸已被試驗性地用於一些抗癌藥物新劑型中。
3.γ-亞麻酸
γ-亞麻酸(γ-lenolenic acid)在1919年由Heidush Kaand Laft於月見草油中發現。目前,富含γ-亞麻酸的月見草油及γ-亞麻酸製品已在營養與醫療方面獲廣泛應用。γ-亞麻酸在臨床上的試驗結果表明其有降血脂作用,對三醯基甘油、膽固醇、p-脂蛋白的下降有效性在60%以上,而且γ-亞麻酸在體內轉變成具有擴張血管作用的PGI2,保持與血栓素A2(TXA2)的平衡,防止血栓形成,從而達到防治心血管疾病的效果。γ-亞麻酸在體內可刺激棕色脂肪組織,促使該組織中線粒體活化,使體內過多熱量能得以釋放,起到防止肥胖症的目的,而且可減輕機體內細胞膜脂質過氧化損害。
4.α-亞麻酸
α-亞麻酸(α-lenolenic acid)最重要的生理功能首先在於它是n-3系列多不飽和脂肪酸的母體,在體內代謝可生成DHA和EPA。由於DHA是腦和視網膜中兩種主要的多不飽和脂肪酸之一,所以,許多動物試驗表明,膳食中α-亞麻酸,特別是在極度或長期缺乏情況下,會出現相應缺乏癥狀,出現視覺循環缺陷與障礙。同時α-亞麻酸的生理功能還表現在對心血管疾病的防治上。Berry和Hirsch在1987年就通過對一組無心臟病或高血壓的中年男子的脂肪組織中的脂肪酸組成分析,指出脂肪組織中α-亞麻酸每增加1%,動脈收縮和舒張壓就降低667Pa。1988年後,Salonen等人觀察到芬蘭男子較低的血壓與α-亞麻酸攝入水平有重要關聯,支持了前述研究結論。我國醫學科學院用富含α-亞麻酸的蘇子油對鼠的高脂血症試驗表明:α-亞麻酸能明顯降低血清中總膽固醇和LDL-膽固醇水平,提高HDL-膽固醇/LDL-膽固醇比值,作用優於安妥明。α-亞麻酸的另一重要功能是增強機體免疫效應。
5.DHA和EPA
從對包括人在內的動物的腦、視網膜和神經組織的分析可以發現,二十二碳六烯酸(doco-sahexaenoic acid.DHA)是其中的主要脂肪酸,是大腦及視網膜的正常發育及功能保持所必需的。其作用機制首先是由於高度的不飽和而形成一個高度流體性的膜環境,除此之外,它還具有不可替代的特殊作用機制。在腦灰色物質和視網膜中,DHA佔2-羥基乙胺磷酸甘油酯中脂肪酸的30%以上。在腦中,DHA和突觸體、突觸小泡、髓磷脂、微粒體、線粒體結合。與花生四烯酸相比,DHA優先結合於視網膜形成三醯基甘油。對貓、猴子等動物的有關DHA與視覺功能的實驗較好地揭示了DHA在視力方面的重要性。而且DHA和EPA(eicosapentaenoic acid,二十碳五烯酸)攝入后,可快速地顯著提高體內這兩種脂肪酸的水平,為其功能的及時發揮提供了保證。因此,在神經系統方面,DHA和EPA被證明具有維持和改善視力,提高記憶、學習等能力,抑制老年痴呆症的生理學效果。
中鏈脂肪酸
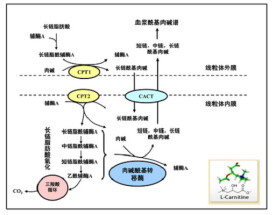
中鏈脂肪酸在體內主要以遊離形式被吸收。由於碳鏈短,中鏈脂肪酸較長鏈脂肪酸水溶性好而容易被胃腸吸收,不會像長鏈脂肪酸在腸內細胞重新酯化。含中鏈脂肪酸的油脂一入口就在舌脂肪酶作用下消化並在胃中繼續水解,舌脂肪酶對富含中鏈脂肪酸的三醯基甘油水解具有專一性,從腸內水解吸收到血液需0.5h,2.5h可達最高峰,是長鏈脂肪酸耗時的一半。中鏈脂肪酸除少量在周圍血液中短期存在外,大部分與白蛋白結合,通過門靜脈系統較快地到達肝臟。在肝臟中,中鏈脂肪酸能迅速通過線粒體雙層膜,在辛醯CoA作用下迅速被醯化,而幾乎不被合成脂肪。醯化產生的過多的乙醯CoA在線粒體胞漿中發生各種代謝作用,其中大部分趨向合成酮體,其生酮作用強於長鏈脂肪酸,而且不受甘油、乳酸鹽、葡萄糖一胰島素等抗生酮物質的影響,在肝臟外組織中,中鏈脂肪酸的代謝作用較少,但在臍帶血中發現C8:0或以下脂肪酸佔15%-20%,這一點也顯示中鏈脂肪酸在胎兒營養中也有生理作用。
主要脂肪的脂肪酸組成及含量
由於中鏈脂肪酸生化代謝相對快速,所以它可作為快速能量來源,特別是對膳食油脂中長鏈脂肪酸難以消化或脂質代謝紊亂的個體,如無膽汁症、胰腺炎、原發性膽汁肝硬化、結腸病、小腸切除、缺乏脂肪酶的早產兒和纖維囊泡症病人等。中鏈脂肪酸的另一重要作用是酮體效應,所有肝外組織可利用它迅速氧化產生大量酮體,手術后病人可利用它來提供熱能,妊娠婦女可通過注射中鏈脂肪酸酯補充胎兒消耗酮體較多的需求。它還能節約慢性病患者肌肉中的肉鹼,改善與敗血症或創傷有關的酮體血症的抑制狀態。此外,中鏈脂肪酸生成的酮體具有麻醉和抗驚厥作用,在臨床上已被用作無抗藥性的癲癇治療藥物。
基本信息
- 中文名
- 脂肪酸
- 外文名
- fatty acids
- 化學品類別有機物
- 有機物
- 構成元素
- 碳、氫、氧構成的化合物
- CAS號
- 67254-79-9
- 常見種類
- 桂酸、豆蔻酸、花生四烯酸
- 物理性狀
- 無色、無臭、無味,相對密度皆小於1
- 生理意義
- 機體主要能量來源之一