異特龍
晚侏羅紀時期最兇猛的恐龍之一
異特龍,又名躍龍或異龍,是蜥臀目獸腳亞目肉食龍下目恐龍的一屬。異特龍是種中型的二足、掠食性恐龍,身長8.5米,最大9.7米,體重1.5~3噸最重3.6噸。它們生存於晚侏羅紀,約1億5500萬年前到1億3500萬年前。自從在1877年被奧塞內爾·查利斯·馬什(Othniel Charles Marsh)命名以來,已有許多的可能種被歸類於異特龍屬,但只有少數被認為是有效種。
異特龍是侏羅紀後期活躍於北美洲、非洲等地的主要肉食性恐龍。第一具異特龍化石是1877年在美國科羅拉多州被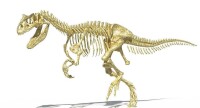 發現的。化石顯示,這種恐龍集猛禽與鱷魚的特性於一身,其學名的意思就是“與眾不同的蜥蜴”。異特龍的化石出土量非常豐富。在目前所發現的該時期的恐龍化石中,異特龍化石佔了將近十分之一,這也為人們研究它提供了豐富的資料。就體形而言,異特龍並不是肉食性恐龍中最大的,但它擁有更適於獵殺的身體結構。首先,它的前肢非常粗壯,上面長有3根指頭,每根指頭上都生有利爪,可以毫不費力地撕開獵物。其次,高大粗壯的後肢能有力地支撐起它的重量,使它行動起來更為敏捷。最後,粗大的尾巴還可以當做鞭子,橫掃任何膽敢進犯的敵人。
發現的。化石顯示,這種恐龍集猛禽與鱷魚的特性於一身,其學名的意思就是“與眾不同的蜥蜴”。異特龍的化石出土量非常豐富。在目前所發現的該時期的恐龍化石中,異特龍化石佔了將近十分之一,這也為人們研究它提供了豐富的資料。就體形而言,異特龍並不是肉食性恐龍中最大的,但它擁有更適於獵殺的身體結構。首先,它的前肢非常粗壯,上面長有3根指頭,每根指頭上都生有利爪,可以毫不費力地撕開獵物。其次,高大粗壯的後肢能有力地支撐起它的重量,使它行動起來更為敏捷。最後,粗大的尾巴還可以當做鞭子,橫掃任何膽敢進犯的敵人。
異特龍
異特龍(Allosaurus)又稱躍龍或異龍,是蜥臀目獸腳亞目恐龍的一屬。異特龍是種中型的二足、掠食性恐龍,身長為9米,最大可達9.7米。它們生存於晚侏羅紀,約1億5500萬年~1億3500萬年前。

異特龍具有大型的頭顱骨,上有大型洞孔,可減輕重量,眼睛上方擁有角冠。它們的頭顱骨是由幾個分開的骨頭組成的,骨頭之間有可活動關節,進食時頜部可先下上張開,然後在左右撐開吞下食物;它們的下頜也可以前後滑動。嘴部擁有70顆大型、銳利、彎曲的牙齒。相較於大型、強壯的後肢,它們的前肢小,手部有三指,指爪大而彎曲,長度為25厘米。尾巴長而重,可平衡身體與頭部。異特龍的骨架和其它獸腳亞目恐龍一般,呈現出類似鳥類的輕巧中空特徵。
異特龍是該時期北美洲莫里遜組最常見的大型掠食動物,並位在食物鏈的頂層。它們可能以其他大型草食性恐龍為食,例如:鳥腳下目、劍龍科、蜥腳下目恐龍。異特龍常被認為採用群體合作方式攻擊蜥腳類恐龍,但很少有證據顯示異特龍具有共同攻擊的社會行為。它們有可能採取伏擊方式攻擊大型獵物,使用上頜來撞擊獵物。
第一個可明確歸類於異特龍的化石,是在1877年由奧塞內爾·查利斯·馬什(Othniel Charles Marsh)所發現。異特龍具有複雜的分類歷史,過去曾有許多種最初被歸類於異特龍,腔軀龍,直到在克利夫蘭勞埃德採石場發現大量的化石后,異特龍才成為常用的名稱,並成為最廣受研究的恐龍之一。
異特龍的化石主要來自於北美洲的莫里遜組,另外在葡萄牙、坦尚尼亞也發現了可能的化石。異特龍的化石是美國猶他州的州化石。由於異特龍是最早被發現的獸腳亞目恐龍之一,所以長期以來吸引了一般大眾的注意,並出現在數個電影與電視節目中。
異特龍是種典型的大型獸腳類恐龍,擁有大型頭顱骨、粗壯的頸部、長尾巴、以及縮短的前肢。脆弱異特龍是最著名的種,平均身長為9米,而最大型的異特龍標本(編號AMNH680)的身長估計為9·7米,體重為3.6噸。在1976年,詹姆斯·麥迪遜(James Madsen)的異特龍專題論文中,他提出異特龍的身長最大值為13米。如同其他的恐龍,異特龍的體重估計值也有爭議,自20世紀80年代以來,成年異特龍的體重估計值,已有1000公斤,以及1010公斤等不同的數據。

異特龍
頭顱骨
在獸腳亞目之中,異特龍的頭顱骨、牙齒與身體的比例適中。葛瑞格利·保羅(Gregory S. Paul)依據一個長度為84.5公分的異特龍頭顱骨,估計該個體的身長為7.9米。每塊前上顎骨各有5顆牙齒,牙齒的橫剖面呈D形,而每塊上顎骨約有14到17顆牙齒;異特龍的牙齒數量與骨頭大小並不呈正比。而每塊齒骨約有14到17顆牙齒,平均數量為16顆。越往嘴部深處,牙齒就越短、狹窄、彎曲。異特龍的牙齒都為鋸齒狀。這些牙齒很容易脫落,所以它們會不斷地生長、替代,並成為常發現的化石。霸王龍嘴裡有60顆牙齒,異特龍牙齒更多,有70顆,而且每顆牙齒像匕首一樣鋒利,所有的牙齒向後彎曲,用於撕開獵物的肉。
異特龍
異特龍的眼睛上方擁有一對角冠,由延伸的淚骨所構成。角冠的形狀與大小隨著個體而不同。鼻骨的上方也有一對低矮的稜脊,並沿者鼻骨,連接到眼睛上的角冠。這些角冠可能覆蓋者角質,並具有不同的功能,例如:替眼睛遮辟陽光、視覺展示物、以及物種內的打鬥行為(問題是它們非常脆弱)。頭顱骨後上方也有一個稜脊,可供肌肉附著,這特徵也可見於暴龍科動物。
淚骨內側有凹處,可能內藏腺體,例如鹽腺。上顎骨內側有凹陷處,發展的比基礎獸腳類恐龍的鼻竇還好,例如角鼻龍與馬什龍;這些凹陷處可能與嗅覺器官有關,例如犁鼻器。異特龍腦殼頂部較薄,可能為促進腦部的體溫調節。
異特龍的頭顱骨是由個別的骨頭所組成,而骨頭之間有可活動關節。例如下顎的前半部與後半部可往外彎曲,增加骨頭間的空隙,因此可以吞下較大的食物。腦殼與額骨之間可能也有類似的關節。
藉助下頜靈活的關節,異特龍的嘴可以張得很大,便於撕咬。這對獵物來說是致命的攻擊。
骨骸
異特龍擁有9節頸椎、14節背椎、5節支撐臀部的薦椎。尾椎的數量不明,可能隨者個體大小而不同;詹姆斯·麥迪遜估計異特龍有接近50節尾椎,而葛瑞格利·保羅認為這個數量過多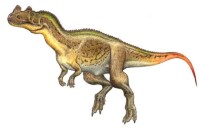 ,提出應該不超過45節。頸椎與前段背椎有中空區域,這種空間也可見於現代鳥類,被認為具有類似鳥類的氣囊系統,使用於呼吸作用上。異特龍的肋骨寬廣,形成桶狀胸腔,與較原始的獸腳類(例如角鼻龍)不同。異特龍也具有腹肋,但不常被發現,可能有稍微的骨化。在一個已公布標本中,這些腹肋被發現生前曾受過傷。有一個叉骨被保存下來,但直到1996年才被確認出來;在一些案例中,叉骨與腹肋則被混淆。主要的臀部骨頭腸骨巨大,恥骨有個明顯的尾端,可能作為肌肉附著處,以及身體躺在地面時的支撐物。在1976年,麥迪遜發現克利夫蘭勞埃德恐龍採石場所發現的異特龍化石,有接近一半個體的兩個恥骨上端,並未互相固定者。由於這個特徵與體型無關,因此麥迪遜認為這是種兩性異形,雌性個體的恥骨上端沒有互相固定著,可使產卵時更為順利。然而,這個理論並未引起進一步的討論。
,提出應該不超過45節。頸椎與前段背椎有中空區域,這種空間也可見於現代鳥類,被認為具有類似鳥類的氣囊系統,使用於呼吸作用上。異特龍的肋骨寬廣,形成桶狀胸腔,與較原始的獸腳類(例如角鼻龍)不同。異特龍也具有腹肋,但不常被發現,可能有稍微的骨化。在一個已公布標本中,這些腹肋被發現生前曾受過傷。有一個叉骨被保存下來,但直到1996年才被確認出來;在一些案例中,叉骨與腹肋則被混淆。主要的臀部骨頭腸骨巨大,恥骨有個明顯的尾端,可能作為肌肉附著處,以及身體躺在地面時的支撐物。在1976年,麥迪遜發現克利夫蘭勞埃德恐龍採石場所發現的異特龍化石,有接近一半個體的兩個恥骨上端,並未互相固定者。由於這個特徵與體型無關,因此麥迪遜認為這是種兩性異形,雌性個體的恥骨上端沒有互相固定著,可使產卵時更為順利。然而,這個理論並未引起進一步的討論。
角鼻龍
腿部

異特龍
智商
通過研究遺骸,我們了解到許多恐龍身體龐大,但這不意味著他們很聰明。馬門溪龍活著的時候約有四五十噸重,而腦子重量只有500克左右。又如劍龍,他的身軀有大象那麼大,而腦子卻小得如約100克的核桃。異特龍也長著龐大的身體,但據推測,他的大腦可能很發達,是侏羅紀時期智商最高的大型肉食恐龍,這也給它們的群居提供了方便。
骨骼特徵
異特龍的顱骨長、寬並且很厚。異特龍還有扁平寬闊的腸骨、末端膨大為足狀的恥骨,股骨比脛骨要長。異特龍的 腳踝長得很高,腳上有鱗片,就像不能飛翔的巨型鳥類的腳,且遠比現存的任何一種鳥類的腳都重。拇指的位置較高,朝向後方,趾爪的核心為骨質,外層則為角質。
腳踝長得很高,腳上有鱗片,就像不能飛翔的巨型鳥類的腳,且遠比現存的任何一種鳥類的腳都重。拇指的位置較高,朝向後方,趾爪的核心為骨質,外層則為角質。
異特龍
尖銳的利齒
異特龍生有70顆邊緣帶鋸齒的牙齒,每顆牙齒都像匕首一樣尖銳,並且都向後彎曲。如果某顆牙齒脫落了或在戰鬥中斷掉了,一顆新的牙齒會很快長出來填補這個空缺。另外,異特龍可以將頜部張得很大,然後再向外擴張,這樣有利於撕碎獵物並且吞食大塊的肉。
最兇猛的恐龍之一
異特龍有強勁的後肢和健壯的尾巴,在捕獵時往往成群出擊。古生物學家曾經發現過一些彎龍的骨骼化石,上面有異特龍的牙齒留下的深深的痕迹,折斷的異特龍牙齒也散布在四周,這表明當時曾發生過一場血腥的捕殺。但異特龍也不是什麼時候都能捕捉到新鮮獵物的,因此,它可能也吃動物的屍體。
異特龍的大量化石几乎涵蓋了所有的年齡層,這使得科學家們可以研究異特龍的生長模式與年齡上限。在科羅拉多州發現的一堆壓碎的蛋化石,可能屬於異特龍,這是目前所發現最年幼的異特龍化石。根據四肢骨頭的組織學分析,異特龍的年齡上限大約為22到28歲,相當於其他大型獸腳類恐龍(例如暴龍)。異特龍的最高生長率大約發生在15歲時,一年可以增加148公斤的體重。已在一個出土於克利夫蘭勞埃德的異特龍脛骨,發現了骨髓骨組織。除了異特龍以外,腱龍與暴龍也發現了骨髓骨。骨髓骨只存在於產卵的雌性鳥類身上,骨髓骨富含鈣,可用來製造蛋殼的。異特龍的骨髓骨組織,顯示該個體是雌性的,而且正在繁衍期中。這個雌性異特龍估計是在10歲時死亡,從此顯示異特龍在完全成長前,就已達到性成熟。
一個具有幾乎完整後肢的幼年異特龍標本,顯示幼年個體的後肢比例較成年個體長,而且後肢下半部(小腿與腳部)長於大腿部分。這些差別顯示年輕異特龍的移動速度較快,並具有不同於成年個體的獵食方式,例如追趕小型獵物,而成年個體則改采伏擊方式捕食大型的獵物。隨者異特龍的成長,它們的大腿骨頭變得更厚、更寬,而橫剖面變得較不圓形,隨者肌肉附著點的改變,肌肉相對更短,腿部的成長減緩。這些改變顯示幼年異特龍的腿部,承受的應力較成年異特龍小,幼年異特龍可能以更規律的速度前進。
異特龍被認為是種主動攻擊的大型掠食者。根據蜥腳類恐龍骨頭上的異特龍齒痕,以及與蜥腳類化石一起發現的零散異特龍牙齒來判斷,異特龍可能以蜥腳類恐龍為獵食對象,或是搜尋它們的屍體為食。另外有明確證據顯示異特龍曾經攻擊過劍龍,例如一個異特龍的尾椎上有個部分痊癒的傷口,這個被刺穿傷口的形狀符合劍龍的尾刺;另外,在一個劍龍的頸部骨板上有個U形的傷口,與異特龍的嘴部形狀符合。在1988年,葛瑞格利·保羅(Gregory Paul)提出異特龍不可能以蜥腳類恐龍為食,除非採取群體方式獵食;因為異特龍的頭部大小屬中型、牙齒相對較小,體型也無法與同時代的大型蜥腳類恐龍相比。另一個可能則是異特龍以幼年蜥腳類恐龍為獵食對象,而不獵食完全成長的蜥腳類恐龍。90年代與2000年的研究可能解答了這個問題。羅伯特·巴克(Robert T. Bakker)將異特龍與一些生存於新生代肉食性哺乳類相比,發現類似 的適應演化,例如:顎部肌肉的縮小、頸部肌肉的增大、以及將顎部左右撐開的能力。雖然異特龍的牙齒並非如這些哺乳類呈軍刀狀,巴克提出了另一種異特龍的攻擊方式:上顎的短牙齒會形成類似鋸子的小型鋸齒表面,可切入獵物肉體。這種型態的顎部可使異特龍採取撕咬方式攻擊大型獵物,消耗獵物的體力。
的適應演化,例如:顎部肌肉的縮小、頸部肌肉的增大、以及將顎部左右撐開的能力。雖然異特龍的牙齒並非如這些哺乳類呈軍刀狀,巴克提出了另一種異特龍的攻擊方式:上顎的短牙齒會形成類似鋸子的小型鋸齒表面,可切入獵物肉體。這種型態的顎部可使異特龍採取撕咬方式攻擊大型獵物,消耗獵物的體力。
異特龍
埃米莉·雷菲爾德(Emily J. Rayfield)等人使用有限元分析,研究了異特龍的頭顱骨,也得到了類似的結果。根據其中的生物力學研究結果,異特龍的頭顱骨非常強壯,咬合力相當大。異特龍的咬合力可以達到3-8噸,而且它們的頭顱可承受約55000牛頓來自於齒列的垂直壓強。這個研究也提出異特龍使用頭部來撞擊獵物,並張開大口、撕咬獵物。這個研究認為異特龍的頭部結構允許它們採取不同的獵食模式來攻擊不同的獵物:它們的頭部較輕型,可攻擊較小、較靈活的鳥腳類恐龍;但頭部有足夠的強度承受撞擊,可允取它們採取伏擊方式攻擊較大型的劍龍科與蜥腳類恐龍。
其他的科學家則對這個研究表示異議,他們認為現存生物中沒有採取撞擊方式獵食的動物,並提出異特龍的頭部有足夠的強度,應可承受獵物掙扎的力量。雷菲爾德等人對異議提出回應,他們承認現存生物中沒有類似異特龍的動物,但異特龍的齒列適合這種攻擊方式,而它們的頭部結構可保護上顎、減低承受力量。另一種可能則是,獸腳類恐龍(例如異特龍)不必費力將獵物殺死,而是從活生的蜥腳類恐龍身上咬下足夠的肉塊,肉塊的大小隻需維持獵食者生存即可。這種獵食方式也使獵物有機會痊癒,而獵食者也可能以類似的方式再度獵食。另外,鳥腳類恐龍是當地最常見的獵物,因此異特龍可能采偷襲方式獵食鳥腳類恐龍,使用前肢抓住獵物,並咬斷獵物喉嚨的氣管,類似今日的大型貓科動物。異特龍的前肢強壯,能夠抓緊獵物,所以這個獵食方式是可能成立的。其他影響進食方式的因素包含:眼睛、前肢、以及後肢。異特龍的頭部形狀將立體視覺限制在20°的範圍內,略小於現代鱷魚。如同鱷魚,這個範圍已足夠異特龍判斷獵物的距離與攻擊時機。相較於其他獸腳類恐龍,異特龍的前肢適合抓住一定距離的獵物,還有將獵物拉近;而指爪的構造顯示它們可用來勾取物體。經推算,異特龍的最高奔跑速度可達每小時30到55公里。
長久以來,半科學文獻與大眾讀物都將異特龍描述成以群體方式獵食,並以蜥腳類恐龍與其他大型恐龍為獵食對象。羅伯特·巴克(Robert T. Bakker)從脫落的牙齒、大型獵物的被咬過骨頭研判,異特龍具有親代養育的社會行為。巴克認為成年異特龍將食物帶到巢穴中,以供幼年異特龍食用,並防止其他肉食性動物找到它們的食物。
研究提出異特龍與其他獸腳類恐龍具有侵略性的物種內行為,而非合作性的行為,如同其他的雙弓動物。一個研究則推論獸腳類恐龍會合作獵食,而非個別獵食;這種行為在脊椎動物中較少見,而現存雙弓動物(包括蜥蜴、鱷魚、鳥類)很少合作獵食。許多現代的掠食性雙弓動物是領域性,會將侵入領地的同類殺死、并吞食它們的屍體;另外當聚集在食物周圍時,它們會將企圖搶先的較小個體殺死。克利夫蘭勞埃德採石場的大量異特龍化石可能源於它們在同類相食時被淹死。這也可以解釋異特龍化石中,幼年與近成年個體所佔的比例較高;因為在現代的鱷魚與科莫多龍中,幼年與近成年個體較少在聚食地點中被殺死。這理論也可解釋巴克所發現的巢穴狀況。有些證據顯示異特龍具有同類相食的行為,例如肋骨碎片上有脫落的同類牙齒、一個肩胛骨上有可能的齒痕、巴克所發現的巢穴中的異特龍骨骸可能遭到同類相食。
一個針對異特龍腦部的電腦斷層掃描,發現它們的腦部與鱷魚和鳥類有較多的共同點。前庭器官的結構顯示它們的頭部保持在幾乎水平的位置,而非朝上或朝下。內耳的結構類似鱷魚,所以異特龍可能容易聽到低頻的聲音,也可以聽到細微的聲音。異特龍的嗅球大,可能適合感覺氣味。
異特龍
異特龍是該時期北美洲莫里遜組最常見的大型掠食者,並位在食物鏈的頂層。它們可能以其他大型草食性恐龍為食,例如:鳥腳目、蜥腳下目恐龍。異特龍經常被認為採用群體合作方式攻擊蜥腳類恐龍,但很少證據顯示異特龍具有共同攻擊的社會行為。它們可能採取伏擊方式攻擊大型獵物,使用上顎來撞擊獵物。
異特龍屬於異特龍科,異特龍科是一個大型獸腳亞目的演化支。奧塞內爾·查利斯·馬什在1878年建立異特龍科;但在70年代以前,異特龍科很少被使用,取而代之的是斑龍科,但斑龍科長期以來被當作大型獸腳類恐龍的“未歸類物種集中地”。在1976年麥迪遜公布他的專題論文以前,跟異特龍相同時期被命名的腔軀龍,常被當作異特龍的同義詞,而且更常被使用。數個研究人員曾在他們的著作中使用斑龍科來取代異特龍科,包含:吉爾摩爾、休尼博士、羅默博士、羅德尼·史提爾、以及亞歷克·沃克。在麥迪遜公布他的具影響力專題論文之後,異特龍科開始普遍使用,但沒有被明確的定義。一些半科學性質的著作將異特龍科使用於不同的大型獸腳類恐龍,通常包含那些比斑龍科更大型、更著名的物種。在獸腳亞目中,常被認為是異特龍近親的物種,包含:印度龍、皮亞尼茲基龍、皮爾逖龍、永川龍、高棘龍、吉蘭泰龍、巧鱷龍、史托龍、以及四川龍。隨者相關恐龍的研究增加,獸腳類恐龍開始變得多樣性。而親緣分支分類法研究則顯示,上述原本被認為是異特龍近親的物種,都不是異特龍科的成員,只有少數屬被歸類於相近的科,例如:高棘龍、永川龍。
異特龍科是肉食龍下目的三個科之一,其他兩個分別為鯊齒龍科與中華盜龍科。在1988年,葛瑞格利·保羅提出異特龍科演化為暴龍科,因此成為並系群;但這個看法已遭到否定,暴龍科已經被歸類為另一群獸腳類支系,虛骨龍類。異特龍科是肉食龍下目中成員最少的一科,在大多數的研究中,除了異特龍屬以外,只有食蜥王龍以及一個發現於法國的未命名異特龍超科恐龍被認為是可能的有效屬。Epanterias是另一個可能的有效屬,但Epanterias與食蜥王龍有可能是異特龍屬的大型個體。最近的研究則保留了食蜥王龍,而將Epanterias歸類於異特龍的一個種。
尚不清楚異特龍屬究竟有多少個種類。自從1988年以來,已有七個種被視為是可能的有效種。例如:模式種脆弱異特龍(A. fragilis)、合異特龍(A. amplexus)、歐洲異特龍(A. europaeus)、尚未有正式敘述的詹氏異特龍(A. jimmadseni)、坦達格魯異特龍(A. tendagurensis),但其中只有少數種普遍被視為有效的種。此外,還有至少10個可疑名稱或未敘述的種曾被歸類於異特龍屬。在基礎堅尾龍類獸腳類恐龍的研究中,只有脆弱異特龍、詹氏異特龍(被列為未命名種)、以及坦達格魯異特龍被承認是可能的有效種;而合異特龍(即為Epanterias)、殘暴異特龍被視為是脆弱異特龍的異名,歐洲異特龍並沒被承認,巨異特龍曾被建立為食蜥王龍。合異特龍、脆弱異特龍、詹氏異特龍、巨異特龍的化石都來自於莫里遜組,莫里遜組散佈於美國的科羅拉多州、蒙大拿州、新墨西哥州、奧克拉荷馬州、南達科他州、猶他州、以及懷俄明州,年代為上侏羅紀的啟莫里階到提通階。在異特龍中,詹氏異特龍化石標本最大。脆弱異特龍是該地層最常發現、最著名的異特龍化石,已發現至少60個個體的化石。自從80年代以來,有爭論懷疑莫里遜組是否存在者另一種異特龍,殘暴異特龍:研究仍維持莫里遜組只有脆弱異特龍的看法,而該地層所發現的不同異特龍化石間的差異,被認為是不同個體間的變化。歐洲異特龍的化石發現於葡萄牙勞爾哈組的波多諾伏,體長只有9米2噸,年代為啟莫里階,可能與脆弱異特龍是同一種動物。坦達格魯異特龍被發現於坦尚尼亞姆特瓦拉區的坦達格魯,年代也為啟莫里階。大部分的研究把坦達格魯異特龍視為異特龍的有效種,但它有可能是一種基礎堅尾龍類,或僅是獸腳亞目的一個疑名。雖然歸類仍不確定,坦達格魯異特龍是種大型獸腳類恐龍,身長可能約9公尺,體重約有2.5公噸。
異特龍的可能異名有腔軀龍、Creosaurus、Epanterias、以及Labrosaurus。而異特龍大部分的種族是根據零碎的化石來命名,許多被認為是破碎異特龍的異名,或者是被錯誤歸類到異特龍屬。其中一個例子是Labrosaurus ferox,是由馬什在1884年所命名,但他是根據一個外形奇怪的部分下顎,而顎部前端的齒列有個明顯的缺口,顎部後段則延展並外翻。後來的研究人員提出這個下顎骨頭具有病狀,應該是該動物生前受傷的痕迹,而後段變形的部分原因為石膏重建的後果。這個化石現在被認為屬於脆弱異特龍。被認為屬於異特龍的化石散佈於全世界,包含澳洲、西伯利亞、以及瑞士,但這些化石被認為是其他的恐龍。
異特龍
早在1869年,科羅拉多州格蘭比郡附近的中央公園d當地居民將一個馬蹄化石交給了費迪南德·范迪威爾·海登(Ferdinand Vandiveer Hayden);該地可能屬於莫里遜組。海登將這標本交給約瑟夫·萊迪(Joseph Leidy),萊迪發現這個“馬蹄”化石其實是半節尾椎骨。約瑟夫·萊迪起初暫時將它歸類為歐洲雜肋龍的一個種(Poicilopleurodon valens)。萊迪後來將它建立為一個新的屬,腔軀龍(Antrodemus)。
異特龍的原型標本(編號YPM 1930)被發現於科羅拉多州卡農城北方的花園公園,由一小群破碎骨頭所構成,包含三節脊椎、一個肋骨碎片、一顆牙齒、一個趾骨、以及右肱骨主幹部分;而右肱骨最常被後來的研究提及。在1877年,奧塞內爾·查利斯·馬什(Othniel Charles Marsh)根據這些化石,把這種生物定名為異特龍,並把其模式種正式命名為脆弱異特龍(Allosaurus fragilis)。種名fragilis來自拉丁語,意即“脆弱的”,是指它們脊椎骨的輕盈特點。
在18世紀後期的化石戰爭期間,馬什與愛德華·德林克·科普(Edward Drinker Cope)之間有過激烈的競爭。兩人曾根據許多零散、相似的化石建立了數個屬,但之後被證明屬於異特龍,使得異特龍的發現與早期研究非常複雜。兩人所建立的名稱包含:馬什命名的Creosaurus(意為“肌肉蜥蜴”)、Labrosaurus(意為“貪吃的蜥蜴”)、以及科普命名的Epanterias(意為“拱起的”)。
在競爭的過程中,科普與馬什並沒有持續地進行他們與他們下屬的挖掘工作。舉例而言,班傑明·福蘭克林·馬奇(Benjamin Franklin Mudge)在科羅拉多州花園公園發現異特龍的原型標本后,馬什便轉向懷俄明州進行新的挖掘工作;而在1883年,M. P. Felch重新開始花園公園的挖掘工作后,卻發現了一個幾乎完整的異特龍化石,以及數個部分骸骨。另外,科普的一個挖掘工人H. F. Hubbell,在1879年於懷俄明州的科摩崖發現了一個異特龍標本(編號AMNH 5753),但他沒有提到化石的完整程度,所以科普從未看過這個化石。在1903年,科普死後數年,這個標本被發現是當時最完整的獸腳類恐龍之一,並在1908年開始展覽。在一個由查爾斯·耐特(Charles R. Knight)所繪製的圖畫中,編號AMNH 5753的異特龍跨越在一隻迷惑龍身上,吞食著迷惑龍的屍體。雖然這是第一次將獸腳類恐龍描繪成站立姿態,但當時並沒有科學證據可以支持。
異特龍
異特龍名稱的重複使得研究變的複雜,並隨者馬什與科普的競爭而惡化。在當時便有一些科學家,例如塞繆爾·溫德爾·威利斯頓(Samuel Wendell Williston),提出有太多相關名稱被重複建立。在1901年,威利斯頓便指出馬什自己也無法分辨出異特龍與Creosaurus的差異。查爾斯·懷特尼·吉爾摩爾(Charles W. Gilmore)在1920年嘗試將這些複雜的名稱整理、歸類。吉爾摩爾認為萊迪用來命名腔軀龍的尾椎,其實跟異特龍的尾椎一樣;因為腔軀龍較早命名,所以取代異特龍。在接下來的50年,腔軀龍取代異特龍,成為正式的名稱,直到詹姆斯·麥迪遜在克利夫蘭勞埃德採石場發現新的異特龍化石,並出腔軀龍的化石材料無法鑒定,發現地點也不清楚,應該採用異特龍這名稱。腔軀龍為非正式的用法,只用在區分吉爾摩爾與麥迪遜兩人重建的不同形態頭顱骨。
早在1927年開始,猶他州艾麥里縣的克利夫蘭勞埃德恐龍採石場便有了零散的發現,威廉·斯托克斯(William J. Stokes)在1945年於《科學》雜誌描述了這個採石場,但直到60年代,才開始了大型的挖掘計畫。在1960年到1965年期間,在接近40個機構的合力挖掘之下,該採石場出土了數千塊骨頭。克利夫蘭勞埃德採石場的著名原因包含:大部分骨頭都屬於脆弱異特龍,例如在2006年,73個出土的恐龍個體中,至少有46個屬於脆弱異特龍;這些化石不呈天然狀態,而且互相混合;將近有10多個科學研究討論了該地點的化石埋葬狀況,形成不同、互相矛盾的解釋。對於該地的成因,包含大群動物深陷在泥濘之中、乾旱導致大群動物困在水窪之中。無論正確的成因為何,當地發現的大量異特龍化石使得科學家可以詳細地研究它們,使得異特龍成為了解最多的獸腳類恐龍之一。該地所出土的異特龍化石几乎包含各種年齡與大小,身長範圍從1公尺到9公尺。
自從麥迪遜的專題論文公布以來,已有許多研究討論異特龍的古生物學以及古生態學。這些研究涵蓋了異特龍的骨骼差異、生長模式、頭顱骨重建、獵食模式、腦部構造、以及群居生活與親代養育的可能性。早期異特龍標本的重新研究、在葡萄牙新發現的化石、以及數個非常完整的標本也有助於對異特龍的了解。
“大艾爾”
在1991年發現的“大艾爾”標本(編號MOR 593),是最著名的異特龍化石之一。“大艾爾”是個相當完整的天然狀態標本,接近95%完整度,身長約8米。“大艾爾”最初是由卡比·希伯(Ki rby Siber)所率領的瑞士團隊發現於懷俄明州的比格霍恩縣,並由懷俄明大學地理博物館與洛磯山博物館共同挖掘出土。因為該化石相當完整,因此取了個昵稱:“大艾爾”。發現“大艾爾”的瑞士團隊後來在同一地點發現另一異特龍化石,並取名為“大艾爾二號”,這具化石是到目前為止,保存狀態最好的異特龍化石。
rby Siber)所率領的瑞士團隊發現於懷俄明州的比格霍恩縣,並由懷俄明大學地理博物館與洛磯山博物館共同挖掘出土。因為該化石相當完整,因此取了個昵稱:“大艾爾”。發現“大艾爾”的瑞士團隊後來在同一地點發現另一異特龍化石,並取名為“大艾爾二號”,這具化石是到目前為止,保存狀態最好的異特龍化石。
異特龍
“大艾爾”的體型小於脆弱異特龍的平均大小,可能是個近成年體,估計只成長到了87%,或是另一較小的種。“大艾爾”由布倫特·布萊特豪普特(Brent Breithaupt)是在1996年所敘述。其中有19個骨頭是斷裂的,呈現出感染的跡象,這可能導致“大艾爾”的死亡。這些感染的骨頭包含五個肋骨、五節脊椎、四個腳部骨頭;數個骨頭則具有骨髓炎。右腳上的感染與傷口可能影響了“大艾爾”的行走,從步態的改變研判,右腳的傷口可能感染了另一隻腳。
異特龍是美國西部莫里遜組地層中最常見的大型獸腳亞目化石,它們位在該地食物鏈的最上層。莫里遜組被認為是半乾旱的環境,具有明顯的雨季和旱季,地形為平坦的泛濫平原。該地層的植被是由針葉樹、樹蕨、蕨類所構成的樹林,以及由蕨類所構成的疏林莽原。
莫里遜組是個富含化石的挖掘地點,該地曾發現綠藻、真菌、苔蘚、木賊、蕨類、蘇鐵、銀杏、以及數科針葉樹等植物化石;其他的動物化石則包含:雙殼綱、蝸牛、輻鰭魚綱、青蛙、蠑螈、烏龜、喙頭目、蜥蜴、陸生與水生的鱷形超目、數種翼龍目、大量的恐龍、以及早期的哺乳類,例如:柱齒獸目、多瘤齒目、對齒獸目、三尖齒獸目。莫里遜組所發現的恐龍則包含了獸腳亞目的角鼻龍、嗜鳥龍、蠻龍,蜥腳下目的迷惑龍、腕龍、圓頂龍、梁龍,鳥臀目的彎龍、橡樹龍、劍龍。
在猶他州的克利夫蘭勞埃德採石場,有一個數量眾多的化石層。這個化石層包含超過10,000個骨頭,大部分屬於異特龍化石,但也有其他恐龍化石,例如劍龍與角鼻龍。如此眾多的動物化石為何集中於同一地點,仍不清楚。而且肉食性恐龍的比例大於草食性恐龍,這種狀況非常的少見。這個化石層被解釋成群體獵食所造成的,但很難證實。另一個可能性是克利夫蘭勞埃德恐龍採石場在過去是個“掠食者陷阱”,類似拉布雷亞瀝青坑,造成大量的掠食者陷入無法掙脫的沉積層中。葡萄牙所發現異特龍的地層,年代屬於晚侏羅紀,被認為環境類似莫里遜組,但受到海洋的影響較多。這個葡萄牙地層也發現了許多莫里遜組的恐龍,主要有異特龍、角鼻龍、蠻龍、迷惑龍,或者是相近物種,例如腕龍的近親葡萄牙巨龍、彎龍的近親龍爪龍。
異特龍與其他侏羅紀獸腳類恐龍共同生存在莫里遜組與葡萄牙,包含:角鼻龍、體型較大的蠻龍,食蜥王龍,依潘龍。這點很不尋常。根據生理結構與化石位置,它們三者似乎擁有不同的生態位。角鼻龍與蠻龍可能活躍於河道附近,角鼻龍身體較細,叫矮,而蠻龍則十分粗壯,擁有威力巨大的爪子和威力無窮的放血牙加碎骨牙,但體 型至少有10-12米+,可使它們躲藏在森林中突襲迷惑龍,腕龍這類大型蜥腳類,有時也會去捕獵劍龍家族的成員;異特龍的腿部較長、速度較快,但它們在森林或矮樹叢中較不靈活,因此可能活躍於乾燥的泛濫平原中。對於角鼻龍的研究比蠻龍還深入。角鼻龍與異特龍的差別在於,它們的頭顱骨較高、較狹窄,而牙齒較大、較寬廣。在一個異特龍的恥骨末端發現了另一個獸腳類恐龍的齒痕,可能是由角鼻龍或蠻龍所留下的,異特龍可能也是其他獸腳類恐龍的食物來源選擇之一,但是恥骨是位於腹部下緣,夾在兩腿之間,處於身體最龐大的地方之一。顯示這隻異特龍是在死後,屍體被其他恐龍所吞食。
型至少有10-12米+,可使它們躲藏在森林中突襲迷惑龍,腕龍這類大型蜥腳類,有時也會去捕獵劍龍家族的成員;異特龍的腿部較長、速度較快,但它們在森林或矮樹叢中較不靈活,因此可能活躍於乾燥的泛濫平原中。對於角鼻龍的研究比蠻龍還深入。角鼻龍與異特龍的差別在於,它們的頭顱骨較高、較狹窄,而牙齒較大、較寬廣。在一個異特龍的恥骨末端發現了另一個獸腳類恐龍的齒痕,可能是由角鼻龍或蠻龍所留下的,異特龍可能也是其他獸腳類恐龍的食物來源選擇之一,但是恥骨是位於腹部下緣,夾在兩腿之間,處於身體最龐大的地方之一。顯示這隻異特龍是在死後,屍體被其他恐龍所吞食。
異特龍

基本信息
- 中文名
- 異特龍
- 拉丁學名
- Allosaurus
- 別名
- 躍龍、異龍
- 分布區域
- 美國、北美洲
- 界
- 動物界
- 門
- 脊索動物門
- 亞門
- 脊椎動物亞門
- 綱
- 蜥形綱
- 亞綱
- 雙孔亞綱
- 總目
- 恐龍總目
- 目
- 蜥臀目
- 亞目
- 獸腳亞目
- 科
- 異特龍科
- 屬
- 異特龍屬
- 種
- 2
- 體長
- 平均約10米,最長12~13米
- 體重
- 約1400~2000千克