革苞菊
菊科旋覆花亞族屬革苞菊屬植物
革苞菊革苞菊,稀有種。多年生草本,僅分佈於內蒙古局部地區海拔1000至1200米的荒漠地帶,為強旱生植物。花期5至6月,本種為蒙古高原植物區系的特有種,對研究亞洲中部植物區系和菊科植物的系統發育有一定的科學意義,革苞菊是一個獨立種。
多年生草本,有粗壯的根狀莖,基部為厚層殘存的枯葉柄所緊密圍裹成徑達6-7厘米的塊狀體;莖基被棉狀污白色厚茸毛,上端有少數稀多數簇生或單生的花莖。花莖不分枝,長2-4厘米,柔弱,徑約2毫米,被白色密茸毛,稍有溝,無葉。葉多數生於莖基上成蓮座狀葉叢,通常長7-15厘米,寬2-4厘米,有基部擴大被長茸毛的葉柄;葉片長圓形,革質,被疏或密的蛛絲狀毛或茸毛,羽狀深裂或淺裂;裂片寬短,有淺齒和生於齒端而長約2-4毫米的硬刺;中脈在下面稍凸起;內層葉較狹。
頭狀花序在莖端單生,下垂,徑達2厘米。總苞倒卵圓形,長約1.5厘米;總苞片3-4層,被蛛絲狀棉毛,外層由較寬長的苞葉組成,革質,綠色,有淺齒和生於齒端的黃色刺;內層較短,線狀披針形,無齒,上部稍紫紅色,頂端有刺。小花多數,花冠管狀,長7-9毫米,干后近白色,頂端褐黃色;裂片卵圓披針形,稍尖,長1.2-1.5毫米。花柱分枝短,卵圓形,頂端稍鈍,下部稍扁;基部膨大。冠毛長5-6毫米,污白色,有不等長而上部稍粗厚的微糙毛。瘦果無毛。
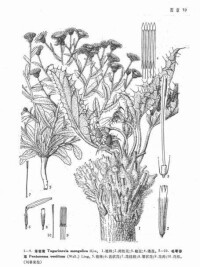
革苞菊
● 小孢子發生和雄配子體發育
小孢子發生:花果期5-6月。革苞菊雄花花藥發育早期,孢原細胞發生於4個角隅處的表皮之下,為多孢原。由孢原細胞分裂分化形成的造孢細胞發育產生小孢子母細胞,小孢子母細胞開始減數分裂時就已分散開來。小孢子母細胞減數分裂Ⅰ不伴隨著胞質分裂,形成四面體型的小孢子四分體,其胞質分裂為同時型。在小孢子孢原、造孢、母細胞中均未見多糖積累,只含可溶性蛋白質。在同一朵花的不同花藥及同一花藥的不同葯室中,小孢子發育是不同步的。可同時見到從小孢子母細胞一直到單核小孢子的不同發育時期。而在同一葯室中小孢子發育時期基本相近。在小孢子四分體中未見多糖積累,只含可溶性蛋白質。包圍4個小孢子的胼胝質物質為PAS正反應。
雄配子體發育:剛從四分體中釋放出來的小孢子的核位於中央,細胞質中只有一些小液泡,壁較薄,內含可溶性蛋白質,無多糖。隨著花粉壁的逐漸增厚,細胞質中小液泡合併成1個大液泡,將小孢子核擠向一邊,即為單核靠邊期。花粉壁在單核後期基本發育完全,此時花粉壁上出現3個萌發孔,其細胞質中除含可溶性蛋白質外,出現了可溶性多糖及少量多糖顆粒。單核花粉經有絲分裂產生二核花粉,其中包含1個生殖核和1個營養核。生殖細胞有絲分裂產生2個紡錘形的精子,與營養核呈品字形排列。花粉成熟散發時,其內壁即從1個萌發孔處開始向外突出,同時2個精子向突出的內壁方向移動,似花粉管開始萌發的跡象。革苞菊的成熟花粉為三細胞,園球形,並具3個萌發孔。其細胞質內含多糖顆粒和可溶性蛋白質,內壁呈PAS正反應,外壁含豐富的蛋白質。同時觀察到,單核小孢子時期產生巨大空花粉現象。觀察了50個花藥的縱切面,巨大空花粉比例約佔10%。巨大空花粉的外壁較薄,細胞質稀少,與正常單核小孢子比較,其體積大出2-3倍。產生這種巨大空花粉的小孢子囊中可包含發育正常的小孢子及絨氈層。
花藥壁發育:在雄花中,初生壁細胞平周分裂,產生內外2層細胞,內層細胞直接發育為絨氈層,外層細胞再分裂1次形成藥室內壁和1層中層。由表皮、葯室內壁、1層中層和絨氈層共同組成花藥的壁。因此,革苞菊的葯壁發育是雙子葉型。中層是短命的,在小孢子發育至四分體時期就開始退化。葯室內壁細胞最終發育為細胞壁條狀加厚的纖維層,並一直保留到花藥成熟開裂。絨氈層細胞多核,在四分體時期尚在原位,單核小孢子後期,絨氈層細胞壁破壞並呈變形蟲狀,原生質周原質團伸入花藥腔中。花粉成熟后,葯壁開裂時只剩不完整的表皮和纖維層。在花藥壁發育過程中,表皮和葯室內壁在花粉成熟前無營養物質積累。花粉成熟后,只在葯室內壁細胞中發現少量可溶性蛋白質。在絨氈層的整個發育過程中,細胞質始終含有大量可溶性蛋白質。而多糖在葯壁發育各階段均未出現在葯壁的各層細胞中。
● 大孢子發生和雌配子體發育
胚珠發育:革苞菊為子房下位,1室,於基部著生1個胚珠。胚珠倒生,單珠被,薄珠心,珠被於大孢子孢原產生時已發育完整。雌花的胚珠一般發育正常,雄花的雌性器官退化或在胚珠靠上部邊緣處分化出似胚囊的幾個細胞,內含一至幾個核。很明顯,這種不正常構造是起不到與正常胚囊一樣的生殖作用的。還發現極少數花的花粉和胚囊均發育正常,與兩性花無區別,但在其胚囊成熟后,卵器一般都呈退化狀態。最終起雄花的作用。在胚珠發育過程中,營養物質的消長可以找到一定規律。其珠心細胞始終含可溶性多糖,珠被絨氈層細胞中則含豐富的可溶性蛋白質。大孢子發育至二核胚囊期,珠被外層細胞,珠柄處和靠近珠孔端的珠被細胞還含有大量多糖顆粒。而到四核胚囊時期,珠被外層細胞多糖顆粒減少,至胚囊成熟時全部消失。此時合點端和珠孔端的珠被細胞則出現可溶性多糖反應。
雌配子體發育:由功能大孢子體積增大並分化形成單核胚囊,單核胚囊又經過3次有絲分裂產生八核胚囊。成熟胚囊包含2個助細胞,1個卵細胞,較早融合的次生核及3個反足細胞。胚囊發育為蓼型。胚囊發育各階段及成熟胚囊中均含可溶性蛋白質,不含多糖。觀察到一定比例的不正常發育的胚囊,如卵細胞退化並在胚囊中出現多核及異形核等現象。
● 雙受精過程的觀察
革苞菊的成熟胚囊為蓼型。受精前,一個助細胞解體,兩極核融合為次生核,卵細胞的核和細胞質偏向於合點端,珠孔端被一個大液泡佔據,使卵表現出明顯極性。花粉管通過珠孔,穿入一個助細胞進入胚囊,釋放內容物,開始雙受精過程。這時,一個精子移向卵細胞,精核靠近卵細胞壁。接著,精核貼在卵細胞壁上。與此同時,另一個精子以同樣的方式靠近次生核的核膜。當一個精核貼在卵細胞壁上時,另一個精核的染色質已經與次生核的核膜融為一體。然後,精子的染色質鬆散,與次生核融合。這時,卵核仁消失,另一精子的染色質進入卵細胞並與卵細胞核融合。由於精核與卵核的融合先於合子的分裂,所以精卵的融合過程屬於有絲分裂前配子配合型。最終產生受精卵和受精極核。受精卵的極性不明顯,在其一側還保留著退化助細胞的痕迹,基部還可見到殘留的花粉管。
● 反足細胞的增生
成熟胚囊的反足細胞為三個,於受精前開始增生。受精后,合子還在休眠時,反足細胞繼續增生。通過連續切片觀察,反足細胞大約可增至15個左右。合子分裂至多細胞原胚時,反足細胞開度始減少。
● 胚和胚柄的發育
合子經過休眠后,第一次分裂為橫向,形成頂細胞和基細胞,基細胞向珠孔處延長,開始形成胚柄吸器。接著頂細胞開始縱分裂,而基細胞則為橫分裂,此時還可見到殘留的花粉管。經過多次分裂后,形成多細胞原胚和不發達的胚柄。由於未得到胚胎髮育後期的材料,所以只觀察到球形胚時期,以後的發育時期還未觀察到。由於受精卵第一次分裂形成的基細胞即開始向珠孔自方向延長,通過以後的分裂,位於胚柄基部的幾個細胞全部延長,伸出珠孔而形成胚柄吸器。
● 胚乳的發育
合子還在休眠時,初生胚乳核開始分裂。第一次分裂就形成細胞壁,接著進行的每一次分裂都形成細胞壁。因此,革苞菊的胚乳發育為細胞型。到多細胞原胚時,已經形成許多胚乳細胞。但百是,其胚囊抄體積較小,胚乳細胞也比較少。
分佈於中國和蒙古南部;在中國分佈於內蒙古中部烏蘭察布盟達爾罕茂明安聯合旗、巴彥淖爾盟烏拉特中旗與后旗、蹬口縣。生於海拔1000-1200米的石質殘丘頂部或礫石質坡地,在中國內蒙古地區,主要生長於於西北部的荒漠草原和荒漠地帶。革苞菊為強旱生植物,生長區的年降水量80-250毫米。在荒漠草原中,主要為小針茅群落的伴生成分,常散生於礫石質坡地的上部。在荒漠中,生長於石質殘丘頂部,可形成局部的革苞菊小居群。
卵葉變種(Tugarinovia mongolicaIljin var.ovatifoliaLing et Y.C.Ma)與原變種的區別在於:葉卵形,側脈3-5對;頭狀花序較大。分佈於中國內蒙古伊克昭盟海渤灣拉僧廟南,低山石礫山坡。
革苞菊僅見於內蒙古烏蘭察布盟北部,巴彥淖爾盟北部、伊克昭盟西部及與蒙古毗鄰的荒漠草原和荒漠地帶。生於海拔1000-1200米的石質殘丘頂部或礫石質坡地。
革苞菊屬是蒙古高原植物區系的特有種屬植物,對研究亞洲中部植物的起源和區系特點有重要價值。

革苞菊
孕蕾期
花蕾早期,在雌花的縱切面上,雌蕊原基剛剛形成時,位於花柱基部的一圈細胞的表皮恢復分裂能力,開始進行垂周分裂。接著表皮以內的幾層細胞也開始分裂分化,從而使花柱基部產生一圈突起。這些細胞較其周圍細胞核大,細胞質濃,為典型的分生組織細胞特點。這個時期的分泌表皮和泌蜜組織還未分化,也沒有多糖和蛋白質的積累,此為蜜腺原基。
花蕾膨大期
雌蕊原基由孢原細胞分化發育至大孢子母細胞階段時,蜜腺原基細胞經過平周和垂周分裂,整個蜜腺體積增大並分化形成分泌表皮和泌蜜組織,並在蜜腺中開始積累少量的澱粉顆粒。氣孔的保衛細胞中積累的澱粉顆粒大而密集。花柱細胞中也看到少量澱粉粒,維管束中則具大量可溶性多糖,未見到蛋白質的積累。從大孢子母細胞減數分裂開始,至二核胚囊形成這一階段,蜜腺繼續長大,澱粉大量增加。
花初放期
雌蕊進一步發育至四核—八核胚囊時,蜜腺中澱粉含量最多,同時期馬虹等:革苞菊花蜜腺的發育解剖學研究出現可溶性蛋白質。這時花柱維管束仍含大量可溶性多糖,其周圍細胞中可見到澱粉粒,蜜腺細胞開始出現較大的液泡。
盛花期
此階段胚囊已發育成熟,是蜜腺泌蜜最旺盛的時期。但在蜜腺中澱粉含量卻開始減少,可溶性蛋白質大量增加,氣孔保衛細胞中含大量澱粉和可溶性多糖,花柱細胞中仍含澱粉,維管束中仍含大量可溶性多糖,維管束中出現少量澱粉粒和可溶性蛋白質。
花敗期
胚囊受精后,蜜腺體積增加到最大,蜜腺細胞中液泡增大,澱粉繼續減少,仍含大量可溶性蛋白質。花柱細胞中澱粉較多,花柱維管束中仍含大量可溶性多糖,蛋白質消失。合子分裂至多細胞原胚期,蜜腺細胞變形,細胞質變稀薄,呈退化趨勢,分泌表皮與泌蜜組織之間出現大的裂隙。蜜腺細胞中含可溶性蛋白較多,澱粉粒較少,氣孔保衛細胞中澱粉粒仍較多。花柱細胞中含少量澱粉粒。
石蠟製片法,在光學顯微鏡下觀察了多糖在革苞菊胚珠發育過程中的消長動態。觀察發現,胚囊發育的不同時期,在珠被、珠柄及子房壁細胞中,多糖含量呈不同程度的變化。而胚囊的整個發育過程始終無多糖積累。受精后,胚和胚乳內均未發現可溶性多糖和儲藏澱粉。珠心細胞中只見到可溶性多糖。認為胚囊發育所需營養是由子房壁通過珠柄到達珠被和胚囊的。

革苞菊
革苞菊為雌雄異株。在雄花中,花藥4室,葯壁發育為雙子葉型,由表皮、葯室內壁,一層中層和絨氈層組成。絨氈層於小孢子四分體時期開始變形,其細胞原生質體向葯室中移動,為變形絨氈層。小孢子孢原為多細胞,小孢子母細胞減數分裂產生四面體型的小孢子四分體。四分體胞質分裂為同時型。成熟花粉3-細胞型。單核期的小孢子出現壁發育不良和巨大及空花粉現象。在雌花中,胚珠是倒生的,單珠被,薄珠心,珠被於孢原期已發育完整。大孢子孢原單細胞。由孢原細胞直接發育形成大孢子母細胞。4個大孢子直線型,蓼型胚囊。於成熟胚囊期觀察到發育異常的胚囊。通過對胚囊發育過程中營養物質消長規律的研究,討論了環境與發育的相關性問題。
列入《中國生物多樣性紅色名錄-高等植物卷》(2013年9月2日)——無危(LC)。
列入《中國國家重點保護植物》——Ⅱ級。
列入《世界自然保護聯盟紅色名錄》(IUCN)中,保護級別為易危(VU)。
由於人為破壞、過度放牧,使其分佈範圍日益縮小,處於瀕危狀態。
保護價值
本種為蒙古高原植物區系的特有種,對研究亞洲中部植物區系和菊科植物的系統發育有一定的科學意義。革苞菊是一個獨立種。因此,革苞菊屬包含了2個種。革苞菊為北阿拉善一東戈壁分佈種,卵葉革苞菊為南阿拉善東部(桌子山-賀蘭山)低山丘陵分佈種,二者形成明顯的替代分佈格局。該屬為阿拉善荒漠特有屬,亦為蒙古高原特有屬。
保護措施
建議在革苞菊分佈區內,採取有效措施,防止過度放牧,並進行引種繁殖試驗。
可以試用種子繁殖。

革苞菊

基本信息
- 中文名
- 革苞菊
- 別名
- 希日-達蘭
- 拉丁學名
- Tugarinovia mongolica
- 界
- 植物界
- 族
- 菜薊族Trib.Cynareae
- 亞族
- 旋覆花亞族
- 門
- 被子植物門Angiospermae[傳統系統]
- 綱
- 木蘭綱Magnoliopsida[Cronquist系統]
- 亞綱
- 菊亞綱Asteridae[Cronquist系統]
- 目
- 菊目Asterales[Cronquist系統]
- 科
- 菊科Compositae[Cronquist系統]
- 亞科
- 飛廉亞科SubFam.Carduoideae
- 屬
- 革苞菊屬Tugarinovia
- 種
- 革苞菊
- 保護級別
- 中國國家二級重點保護植物
- 分布區域
- 內蒙古自治區
- 命名者及年代
- Iljin,1928