體細胞遺傳學
體細胞遺傳學
目錄
以高等生物的體細胞為實驗材料,採用細胞離體培養、細胞融合和遺傳物質在細胞間轉移等方法,研究真核細胞的基因結構功能及其表達規律等的遺傳學分支學科。它可以為植物育種提供新的方法,並且是人類遺傳性疾病基因治療的理論基礎。
高等生物的遺傳學研究一般都通過分析遺傳性狀在有性生殖子代中的分佈和出現頻率來進行。可是高等生物的生殖周期長,子代個體數目少,對於人類來講則又不能在嚴格的實驗條件下進行雜交實驗,所以給研究帶來了一定的困難。但是作為高等生物個體生命活動的基本單位的每一體細胞一般都包含著全套基因組,因此將體細胞在離體條件下培養,使之像原生動物或細菌一樣分裂、增殖,便可將微生物遺傳學的一些方法應用於高等生物的體細胞研究。例如可以分離體細胞並培養成為純種──克隆(又稱無性繁殖系);可以定量研究各種理化因素對高等生物體細胞的作用,包括突變的誘發;可以克服生殖隔離而實現不同種(如人和鼠)之間甚至不同界(如人和大豆)之間的體細胞融合而獲得細胞雜種;通過體細胞克隆的擴增,在短期內獲得數目眾多的子代細胞從而有效地分析特定性狀的遺傳規律。在植物細胞中更可藉助於植物細胞的全能性使單細胞克隆和融合細胞株分化成為完整的新植株,以便研究雜種細胞的基因表達,分析和比較無性雜種後代的遺傳變異規律,並進行育種。
簡史 1907年美國學者R.G.哈里森第一次把神經細胞在體外培養成活。1956年美國學者T.T.帕克使單個哺乳動物體細胞在體外培養的條件下分裂增殖成為克隆,首次提供了用微生物學方法在嚴格控制的條件下進行體細胞遺傳學研究的材料,簡化了體外獲得高等動物體細胞克隆的程序,把體細胞遺傳學的研究推進到一個新的階段。60年代初法國學者G.巴斯基等發現體外培養的小鼠細胞能自發融合,以後,日本學者岡田善雄又發現經紫外線滅活的仙台病毒可促進細胞融合。在此後的幾年裡體細胞遺傳學的研究相繼取得三項重大進展:①引進了不能合成某些酶的隱性突變細胞株作為細胞融合的親本;在混合兩種不同的隱性突變細胞時,只有發生了融合的細胞才能由於兩個突變型細胞的功能上的互補(見互補作用)而呈現出野生型特性,進而可用各種選擇技術把這些融合細胞從大量未融合的細胞群體中篩選出來;②在獲得包含不同物種的細胞核的種間異核細胞以後,又獲得了體外培養條件下能分裂增殖的合核細胞,即由不同物種的細胞核融合成為一個細胞核的雜種細胞;③發現兩個物種的細胞融合后,雜種細胞在分裂過程中總是連續地排斥其中一個物種的染色體。例如小鼠細胞與人體細胞形成的雜種細胞在分裂過程中傾向於不斷地排斥人的染色體。在上述三個發現的基礎上,迅速地開展了人的基因定位工作、正常體細胞與腫瘤細胞的雜種細胞的致瘤性的研究以及細胞核、細胞質與某些染色體之間的關係的研究。1975年,阿根廷學者C.米爾斯坦等把能產生大量異常免疫球蛋白的小鼠骨髓瘤細胞與經過特定抗原免疫的小鼠脾臟淋巴細胞進行體細胞雜交,從雜種骨髓瘤細胞中得到了產生單克隆抗體的細胞株,大大地提高了抗體的純度,對免疫學和醫學的研究產生了重大影響。
此外,還採用各種方法在體細胞之間轉移遺傳物質,以便觀察外源基因在宿主細胞中的命運。1967年第一次以微生物轉化方法將田鼠黑色素瘤細胞中的 DNA轉化體外培養的非黑色素細胞,使之成為能產生黑色素的細胞並增殖為克隆。70年代中又陸續發展出一些新技術,如製備出微細胞、脂質體和血影細胞等載體,把若干條染色體、染色體片段或長度不等的 DNA分子引入受體細胞;或者通過顯微注射把 DNA分子直接注入受體細胞的核內,最後使這些引入的外源遺傳物質在受體細胞中表達,從而大大推進了有關真核生物的基因結構和功能以及基因調控方面的研究。
植物的體細胞遺傳學研究工作是在植物組織培養的基礎上發展起來的。1934年英國學者P.R.懷特以番茄根為材料建成了第一個能活躍生長的細胞無性繁殖系。以後的發展主要是關於培養物的組織分化和細胞融合兩個方面。1956年R.A.米勒發現了激動素,並且在含有一定濃度的激動素和生長素的培養基上使離體培養的組織發生器官分化。到目前為止,已有200多種、屬植物組織培養中的細胞,相繼被誘導分化為植株。
60年代初期E.C.科金等應用纖維素酶分離植物原生質體獲得成功。1972年P.S.卡爾森通過選擇性篩選方法,獲得由無性雜種細胞分化而成的雙二倍體煙草植株。70年代以來在對突變型細胞株的篩選,外源遺傳物質導入和高等植物細胞中質粒的研究等方面也和動物體細胞遺傳學一樣得到了廣泛地開展。特別是因為植物細胞具有全能性,能夠由單個細胞長成植株,所以植物體細胞遺傳學研究除了在闡明植物的器官分化及形態建成等方面的價值外,也對育種工作具有重要意義。
研究方法 細胞融合 用滅活的病毒(如仙台病毒)或化學藥物(如聚乙二醇、葡聚糖)等處理細胞,可以促使細胞間發生融合而獲得不同的細胞雜種。例如含有兩個以上同種細胞核的同型核細胞、含有兩個以上異種細胞核的異核細胞、含有由同種(或異種)細胞核融合成一個細胞核的種內(或種間)合核細胞、由不含細胞核的胞質體和完整細胞融合成的胞質雜種細胞以及由一種細胞的胞質體同另一種細胞的不含細胞質的核體融合成的重組細胞等。這些通過無性過程獲得的細胞雜種除可以用以研究細胞核和細胞質對遺傳信息的傳遞和表達所起的作用以及腫瘤發生的機理等,還可在植物中用來克服遠緣雜交的困難。
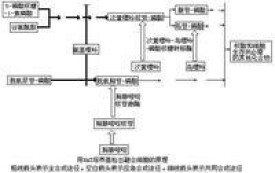
融合細胞的選擇 細胞融合和基因突變同屬稀有事件,所以必須具備特定的選擇方法才能從大量沒有融合的細胞中分離已經融合的細胞。在動物細胞中最早出現而且應用最廣泛的是HAT選擇法。在這一方法中,一個親本細胞株為次黃嘌呤-鳥嘌呤-磷酸核糖轉移酶缺陷型(HGPRT-);另一個親本細胞株為胸腺嘧啶核苷激酶缺陷型(TK-),在含有次黃嘌呤(H)、氨基蝶呤(A)、胸腺嘧啶核苷(T)的HAT選擇培養液中上述親本細胞都無法生存,只有融合以後的雜種細胞才能生長,因此可以有效地選擇出雜種細胞。 HAT選擇法的原理是因為核酸合成有兩條途徑:①全合成途徑,從一些小分子物質先合成嘌呤、嘧啶,最後合成核酸;②應急途徑,通過HGPRT的催化作用把次黃嘌呤轉化成次黃嘌呤核苷-磷酸(IMP),通過TK的催化把胸腺嘧啶核苷轉化成脫氧胸腺嘧啶核苷-磷酸(dTMP),再進一步合成核酸。 HAT培養液中的氨基蝶呤(A)能阻斷全合成途徑,所以兩個親本細胞都不能在HAT培養基中生長,而只有經細胞融合后得到的雜種細胞同時具有了HGPRT和 TK的酶活性才能利用培養液中的次黃嘌呤(H)和胸腺嘧啶核苷(T)通過應急途徑合成核酸(見圖)。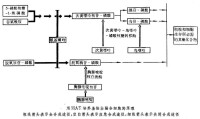 同理,任何其他方法,只要具備能使兩種親本細胞不能單獨生存而只有融合以後才能生存的條件,都可以用來進行融合細胞的選擇。
同理,任何其他方法,只要具備能使兩種親本細胞不能單獨生存而只有融合以後才能生存的條件,都可以用來進行融合細胞的選擇。
體細胞遺傳學
排除或保留某一親本染色體的方法 對於都是長期傳代培養的小鼠和大鼠細胞株的融合細胞來講,在傳代過程中被不斷排斥的是大鼠的染色體;小鼠和人的融合細胞中被排斥的則是人的染色體。可是如果用人體細胞株的細胞與未經長期傳代培養的小鼠細胞進行融合,則融合細胞首先排斥小鼠染色體。在植物中,培養九個月以後的大豆和煙草雜種細胞株中大豆染色體全部保留,煙草染色體則被排除了一半。
根據染色體上某些基因的特性,可以得到選擇性地排除某一染色體的雜種細胞,例如在人的第5號染色體上有一個白喉毒素受體蛋白基因,它使人體的雜種細胞對白喉毒素敏感。小鼠細胞中沒有這一基因,所以能抗這種毒素。把人體和小鼠細胞的融合細胞培養在含有白喉毒素的培養液中,就能選擇性地除去帶有人體第5號染色體的雜種細胞。相反地也可以使融合細胞選擇性地保留某一染色體。例如HGPRT基因位於人的X染色體上,所以能在HAT培養液中生長的融合細胞必定保留有人的X染色體。
染色體基因或細胞核的轉移 把遺傳物質引入某種生物的細胞是體細胞遺傳學研究中的常用手段。外源遺傳物質直接轉化受體細胞的效率不高。把待轉移的遺傳物質用病毒、血影細胞、微細胞、脂質體等裝載后則能提高引入受體細胞的效率。此外,也可以用極細的玻璃管把 DNA直接注射到受體細胞的細胞核中。對於除去細胞壁后的植物細胞的原生質體來講,也可以用上述種種方法引入DNA分子、噬菌體顆粒和細胞器等。除了轉移染色體或染色體片段以外,有時需要轉移整個細胞核,顯微注射也是轉移細胞核的常用方法。
應用 應用細胞融合、染色體鑒定、生化鑒定、免疫學鑒定等技術,已經建立了許多種基因定位方法,使人的基因定位的研究取得了快速的進展。例如,可利用中國倉鼠的細胞和人的體細胞融合的雜種細胞在傳代培養過程中不斷排斥人的染色體的現象來進行基因定位:如發現雜種細胞中人的9號染色體被排斥后便失去ABO血型抗原,就可以確定ABO血型抗原基因是在9號染色體上等。研究腫瘤細胞融合形成的雜種細胞的致瘤性的變化可以為了解正常細胞的癌化和腫瘤細胞的逆轉提供重要的線索。體細胞遺傳學方法還可應用於腫瘤(例如著色性干皮病)發生機理的研究(見DNA損傷修復、互補作用)。另外,體細胞培養的方法還可應用於人類遺傳性疾病的預防(見染色體病、先天性代謝缺陷)。
遺傳物質引入受體細胞后,分析外源基因在受體細胞中是否表達以及了解基因表達時所需的條件是研究真核細胞基因調控的有效途徑。此外,研究細胞核和細胞質之間的相互關係以及細胞質對核基因表達的作用,也都是體細胞遺傳學研究的重要內容。
體細胞遺傳學原理在育種工作中的應用目前還只限於植物。在藥用植物黃連的細胞株中曾選到總生物鹼含量高達乾重的15%(其中小櫱鹼含量高達 8%)的細胞株,這些細胞株可以用來進行小櫱鹼的發酵生產。通過誘變處理還曾從玉米的體細胞中選得抗長蠕孢霉的細胞株並育成抗性品系。通過原生質體融合還可以獲得難以通過有性雜交取得的遠緣雜交品系,例如馬鈴薯和番茄、煙草和矮牽牛的雜交品系。隨著研究工作的深入,將會在生產實際中取得更多成果。
參考書目
N.R.Ringertz,R.E.Savage,Cell Hybrids,Academic Press, New York,1976.