下丘腦
下丘腦
下丘腦又稱丘腦下部。位於大腦腹面、丘腦的下方,是調節內臟活動和內分泌活動的較高級神經中樞所在。通常將下丘腦從前向後分為三個區:視上部位於視交叉上方,由視上核和室旁核所組成;結節部位於漏斗的後方;乳頭部位於乳頭體。下丘腦位於丘腦下鉤的下方,構成第三腦室的下壁,界限不甚分明,向下延伸與垂體柄相連。下丘腦面積雖小,但接受很多神經衝動,故為內分泌系統和神經系統的中心。它們能調節垂體前葉功能,合成神經垂體激素及控制自主神經和植物神經功能。
下丘腦的神經分泌物是通過門脈流如垂體前葉的,有的激發垂體前葉的釋放,稱釋放激素(RH);有的抑制垂體前葉激素的釋放,稱抑制激素(IH)。抑制的促激素釋放或抑制激素有:促甲狀腺激素釋放素(TRH)、促腎上腺皮質激素釋放釋放激素(cRH)、促卵泡生成激素釋放激素(FSH-RH)、促黃體生成激素(LH-RH)、生長激素釋放激(GRH)、生長激素抑制激素(GIH或S.S.)、泌乳激素釋放激(PRH)、黑色細胞刺激素一直激素(MRIH)及黑色細胞刺激素釋放激素(MRH)等十種。下丘腦分泌的釋放抑制激素、垂體分泌的促激素和靶腺合成的激素,形成一個激素網,調節著集體的許多活動。
下丘腦與神經垂體和腺垂體的聯繫非常密切,如視上核和室旁核的神經元軸突延伸終止於神經垂體,形成下丘腦-垂體束。在下丘腦與腺垂體之間通過垂體門脈系統發生功能聯繫。下丘腦的一些神經元既能分泌激素(神經激素),具有內分泌細胞的作用,又保持典型神經細胞的功能。它們可將從大腦或中樞神經系統其他部位傳來的神經信息,轉變為激素的信息,起著換能神經元的作用,從而以下丘腦為樞紐,把神經調節與體液調節緊密聯繫起來。所以,下丘腦與垂體一起組成下丘腦-垂體功能單位。
凡是能分泌神通肽或肽類激素的神經分泌細胞稱為肽能神經元。下丘腦的肽能神經元主要豐碩盱視上核、室旁核與促垂體核團。促垂體區核團位於下丘腦的內側基底部,主要包括正中隆起、弓狀核、腹內側核、視交叉上核以及室周核等,多屬於小細胞肽能神經元,其軸突投射到正中隆起,軸突末梢與垂體門脈系統的第一級毛細血管風接觸,可將下丘腦調節肽釋放進入門脈系統,從而調節垂體的分泌活動。
下丘腦位於丘腦溝以下,構成第三腦室的下壁,界限不甚分明,向下延伸與垂體柄相連。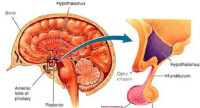 下丘腦面積雖小,但接受很多神經衝動,故為內分泌系統和神經系統的中心。它們能調節垂體前葉功能,合成神經垂體激素及控制自主神經和植物神經功能。下丘腦的神經分泌物是通過門脈流入垂體前葉的,有的激發垂體前葉的釋放,稱釋放激素(RH);有的抑制垂體前葉激素的釋放,稱抑制激素(IH)。釋放的促激素釋放或抑制激素有:促甲狀腺激素釋放素(TRH)、促腎上腺皮質激素釋放激素(cRH)、促卵泡生成激素釋放激素(FSH-RH)、促黃體生成激素釋放激素(LH-RH)、生長激素釋放激素(GRH)、生長激素抑制激素(GIH或S.S.)、泌乳激素釋放激素(PRH)、黑色細胞刺激素抑制激素(MRIH)及黑色細胞刺激素釋放激素(MRH)等十種。下丘腦分泌的釋放抑制激素、垂體分泌的促激素和靶腺合成的激素,形成一個激素網,調節著機體的許多活動。
下丘腦面積雖小,但接受很多神經衝動,故為內分泌系統和神經系統的中心。它們能調節垂體前葉功能,合成神經垂體激素及控制自主神經和植物神經功能。下丘腦的神經分泌物是通過門脈流入垂體前葉的,有的激發垂體前葉的釋放,稱釋放激素(RH);有的抑制垂體前葉激素的釋放,稱抑制激素(IH)。釋放的促激素釋放或抑制激素有:促甲狀腺激素釋放素(TRH)、促腎上腺皮質激素釋放激素(cRH)、促卵泡生成激素釋放激素(FSH-RH)、促黃體生成激素釋放激素(LH-RH)、生長激素釋放激素(GRH)、生長激素抑制激素(GIH或S.S.)、泌乳激素釋放激素(PRH)、黑色細胞刺激素抑制激素(MRIH)及黑色細胞刺激素釋放激素(MRH)等十種。下丘腦分泌的釋放抑制激素、垂體分泌的促激素和靶腺合成的激素,形成一個激素網,調節著機體的許多活動。
下丘腦
下丘腦能通過下述三種途徑對機體進行調節: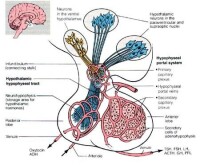 ①由下丘腦核發出的下行傳導束到達腦幹和脊髓的植物性神經中樞,再通過植物性神經調節內臟活動;②下丘腦的視上核和室旁核發出的纖維構成下丘腦——垂體束到達神經垂體,兩核分泌的加壓素(抗利尿激素)和催產素沿著此束流到神經垂體內貯存,在神經調節下釋放入血液循環;③下丘腦分泌多種多肽類神經激素對腺垂體的分泌起特異性刺激作用或抑制作用,稱為釋放激素或抑制釋放激素。下丘腦通過上述算途徑,調節人體的體溫、攝食、水平衡、血壓、內分泌和情緒反應等重要生理過程。如損毀雙側下丘腦的外側區,動物即拒食拒飲而死亡;損毀雙側腹內側區,則攝食量大增引起肥胖。體溫調節的高級中樞位於下丘腦,下丘腦前部受損,動物或人的散熱機制就失控,失去在熱環境中調節體溫的功能;如後部同時受損傷,則產熱、散熱的反應都將喪失,體溫將類似變溫動物。損壞下丘腦可導致煩渴與多尿,說明它對水平衡的調節有關。
①由下丘腦核發出的下行傳導束到達腦幹和脊髓的植物性神經中樞,再通過植物性神經調節內臟活動;②下丘腦的視上核和室旁核發出的纖維構成下丘腦——垂體束到達神經垂體,兩核分泌的加壓素(抗利尿激素)和催產素沿著此束流到神經垂體內貯存,在神經調節下釋放入血液循環;③下丘腦分泌多種多肽類神經激素對腺垂體的分泌起特異性刺激作用或抑制作用,稱為釋放激素或抑制釋放激素。下丘腦通過上述算途徑,調節人體的體溫、攝食、水平衡、血壓、內分泌和情緒反應等重要生理過程。如損毀雙側下丘腦的外側區,動物即拒食拒飲而死亡;損毀雙側腹內側區,則攝食量大增引起肥胖。體溫調節的高級中樞位於下丘腦,下丘腦前部受損,動物或人的散熱機制就失控,失去在熱環境中調節體溫的功能;如後部同時受損傷,則產熱、散熱的反應都將喪失,體溫將類似變溫動物。損壞下丘腦可導致煩渴與多尿,說明它對水平衡的調節有關。
下丘腦
下丘腦對情緒反應有影響,切除大腦皮層而保留下丘腦的動物,可自發產生或輕微刺激就能引起假怒的情緒表現,如猛甩尾巴、豎毛、張牙舞爪、掙扎、瞳孔擴大、呼吸加快、血壓升高等。去除下丘腦的動物,只能零星地表現出上述部分反應。刺激貓下丘腦前區,會引起低頭、耳向後倒、拱背吼叫、肌肉緊張等恐懼反應。這都說明下丘腦與情緒反應關係密切。上述下丘腦的種種功能,有許多是和邊緣系統其他部位的活動密切相關的,而並非下丘腦所獨立完成的。
下丘腦是大腦皮層下調節內臟活動的高級中樞,它把內臟活動與其他生理活動聯繫起來,調節著體溫、攝食、水平衡和內分泌腺活動等重要的生理功能。
(一)體溫調節動物實驗中觀察到,在下丘腦以下橫切腦幹后,其體溫就不能保持相對穩定;若在間腦以上切除大腦後,體溫調節仍能維持相對穩定。現已肯定,體溫調節中樞在下丘腦;下丘腦前部是溫度敏感神經元的所在部位,它們感受著體內溫度的變化;下丘腦後部是體溫調節的整合部位,能調整機體的產熱和散熱過程,以保持體溫穩定於一定水平。
(二)攝食行為調節用埋藏電極刺激清醒動物下丘腦外側區,則引致動物多食,而破壞此區后,則動物拒食;電刺激下丘腦腹內側核則動物拒食,破壞此核后,則動物食慾增大而逐漸肥胖。由此認為,下丘腦外側區存在攝食中樞,而腹內側核存在所謂飽中樞,後者可以抑制前者的活動。用微電極分別記錄下丘腦外側區和腹內側核的神經元放電,觀察到動物在飢餓情況下,前者放電頻率較高而後者放電頻率較低;靜脈注入葡萄糖后,則前者放電頻率減少而後者放電頻率增多。說明攝食中樞與飽中樞的神經元活動具有相互制約的關係,而且這些神經元對血糖敏感,血糖水平的高低可能調節著攝食中樞和飽中樞的活動。
(三)水平衡調節水平衡包括水的攝入與排出兩個方面,人體通過渴感引起攝水,而排水則主要取決於腎臟的活動。損壞下丘腦可引致煩渴與多尿,說明下丘腦對水的攝入與排出均有關係。下丘腦內控制攝水的區域與上述攝食中樞極為靠近。破壞下丘腦外側區后,動物除拒食外,飲水也明顯減少;刺激下丘腦外側區某些部位,則可引致動物飲水增多。下丘腦控制排水的功能是通過改變抗利尿激素的分泌來完成的。下丘腦內存在著滲透壓感受器,它能感受血液的晶體滲透壓變化來調節抗利尿激素的分泌;滲透壓感受器和抗利尿激素合成的神經元均在視上核和室旁核內。一般認為,下丘腦控制攝水的區域與控制抗利尿激素分泌的核團在功能上是有聯繫的,兩者協同調節著水平衡。
(四)對腺垂體激素分泌的調節下丘腦的神經分泌小細胞能合成調節腺垂體激素分泌的肽類化學物質,稱為下丘腦調節肽。這些調節肽在合成后即經軸突運輸並分泌到正中隆起,由此經垂體門脈系統到達腺垂體,促進或抑制某種腺垂體激素的分泌。下丘腦調節肽已知的有九種:促甲狀腺激素釋放激素、促性腺素釋放激素、生長素釋放抑制激素、生長素釋放激素、促腎上腺皮質激素釋放激素、促黑素細胞激素釋放因子、促黑色細胞激素釋放抑制因子,催乳素釋放因子、催乳素釋放抑制因子。
(五)對情緒反應的影響下丘腦內存在所謂防禦反應區,它主要位於下丘腦近中線兩旁的腹內側區。在動物麻醉條件下,電刺激該區可獲得骨骼肌的舒血管效應(通過交感膽鹼能舒血管纖維),同時伴有血壓上升、皮膚及小腸血管收縮、心率加速和其他交感神經性反應。在動物清醒條件下,電刺激該區還可出現防禦性行為。在人類,下丘腦的疾病也往往伴隨著不正常的情緒反應。
(六)對生物節律的控制下丘腦視交叉上核的神經元具有日周期節律活動,這個核團是體內日周期節律活動的控制中心。破壞動物的視交叉上核,原有的一些日周期節律性活動,如飲水、排尿等的日周期即喪失。視交叉上核可能通過視網膜-視交叉上核束,來感受外界環境光暗信號的變化,使機體的生物節律與環境的光暗變化同步起來;如果這條神經通路被切斷,視交叉上核的節律活動就不再能與外界環境的光暗變化發生同步。
下丘腦能神經元與來自其他部位的神經纖維有廣泛的突觸聯繫,其神經遞質比較複雜,可分為兩大類:一類遞質是肽類物質,如腦啡肽、β-內啡肽、神經降壓素、P物質、血管活性腸肽及膽囊收縮素等;另一類遞質是單胺類物質,主要有多巴胺(DA)、去甲腎上腺素(NE)與5-羥色胺(5-HT)。
組織化學研究表明,三種單受類遞質的濃度,以下丘腦“促垂體區”正中隆起附近最高。單胺能神經元可直接與釋放下丘腦調節肽的肽能神經元發生突觸聯繫,也可以通過多突觸發生聯繫。單胺能神經元通過釋放單胺類遞質,調節肽能神經元的活動。下丘腦單受能神經元的活動不斷受中樞神經系統其他部位的影響,所以它們對下丘腦調節肽分泌的調節作用比較複雜。阿片肽對下丘腦調節肽的釋放有明顯的影響。例如,給人注射腦啡肽或β-內啡肽可抑制CRH的釋放,從布使ACTH分泌減少,而納洛酮則有促進CRH釋放的作用;注射腦啡肽或β-內啡肽可刺激下丘腦釋放TRH和GHRH,使腺垂體分泌TSH與GH增加,而對下丘腦的GnRH釋放則明顯的抑制作用。
簡介
下丘腦促垂體區肽能神經元分泌的肽類激素,主要作用是調節腺垂體的活動,因此稱為下丘腦調節肽(hypothalamusregulatorypeptideHRP)。從下丘腦組織提取肽類激素獲得成功,並已能人工合成。1968年Guillemin實驗室從30萬隻羊的下丘腦中成功地分離出幾毫克的促甲狀腺激素釋放激素(TRH),並在一年後確定其化學結構為三肽。在這一生成成果鼓舞下,Schally實驗室致力於促性腺激素釋放激素(GnRH)的提取工作。1971年他們從16萬頭豬的下丘腦中提純出GnRH,又經過6年的研究,闡明其化學結構為十肽。此後,生長素釋放抑制激素(GHRIH)、促腎上腺皮質激素釋放激素(CRH)與生長素釋放激素(GHRH)相繼分離成功,並確定了化學結構,此外,還有四種對腺垂體催乳素和促黑激素的分泌起促進或抑制作用的激素,因尚未弄清其化學結構,所以暫稱因子。
下丘腦調節肽除調節腺垂體功能外,它們幾乎都具有垂體外作用,而且它們也不僅僅在下丘腦“促垂體區”產生,還可以大中樞神經系統其他部位及許多組織中找到它們蹤跡,使人們更加廣泛深入地研究他們的作用。
(一)促甲狀腺激素釋放激素
促甲狀腺激素釋放激素(thyrotropin-releasinghormoneTRH)是三肽,其化學結構為:(焦)谷-組-脯-NH2TRH主要作用於腺垂體促進促甲狀腺激素(TSH)釋放,血中T4和T3隨TSH濃度上升而增加。給人和動物靜脈注射TRH(1mg),1-2min內血漿TSH濃度便開始增加,10-20min達高峰,TSH的含量可增加20倍。腺垂體的促甲狀腺激素細胞的膜上的TRH受體,與TRH結合后,通過Ca2介導引起TSH釋放,因此IP3-DG系統可能是TRH發揮作用的重要途徑。TRH除了刺激腺垂體釋放TSH外,也促進催乳互的釋放,但TRH是否參與催乳素分泌的生理調節,尚不能肯定。下丘腦存在大量的TRH神經元,它們主要分佈於下丘腦中間基底部,如損毀下丘腦的這個區域則引起TRH分泌減少。TRH神經元合成的TRH通過軸漿運輸至軸突末梢貯存,延伸到正中隆起初級毛細血管周圍的軸突末梢在適當刺激作用下,釋放TRH並進入垂體門脈系統運送到腺垂體,促進TRH釋放。另外,在第三腦室周圍尤其是底部排列有形如杯狀的腦室膜細胞(tanycyte),其形態特點與典型的腦室膜細胞有所不同,其胞體細長,一端面向腦室腔,其邊界上無纖毛而有突起,另一端則延伸至正中隆起的毛細血管周圍。在這些細胞內含有大量的TRH與GnRH等肽類激素。下丘腦特別是室周核釋放的TRH或GnRH進入第三腦室的腦脊液中,可被腦室膜細胞攝入,再轉幸福至正中隆起附近釋放,然後進入垂體門脈系統。除了下丘腦有較多的TRH外,在下丘腦以外的中樞神經部位,如大腦和脊髓,也發現有TRH存在,其作用可能與神經信息傳遞有關。
(二)促性腺激素釋放激素
促性腺激素釋放激素(gonadotropin-releasinghormoneGnRHLRH)是十肽激素,其化學結構為:(焦)谷-組-色-絲-酪-甘-亮-精-脯-甘-NH2GnRH促進性腺垂體合成與釋放促性腺激素。當機體靜脈注射100mgGnRH,10min后血中黃體生成素(LH)與卵泡刺激素(FSH)濃度明顯增加,但以LH的增加更為顯著。在體外腺垂體組織培養系統中加入GnRH,亦能引起LH與FSH分泌增加,如果先用GnRH抗血清處理后,再給予GnRH,則可減弱或消除GnRH的效應。下丘腦釋放GnRH的特脈衝式釋放,因而造成血中LH與FSH濃度也呈現脈衝式波動。從恆河猴垂體門脈血管收集的血樣測定GnRH含量,呈現陣發性時高時低的現象,每隔1-2h波動一次。在大鼠,GnRH每隔20-30min釋放一次,如果給大鼠注射抗GnRH血清,則血中LH與FSH濃度的脈衝式波動消失,說明血中LH與FSH的脈衝式波動是由下丘腦GnRH脈衝式釋放決定的。用青春期前的幼猴實驗表明,破壞產生GnRH的弓狀核后,連續滴注外源的GnRH並不能誘發青春期的出現,只有按照內源GnRH所表現的脈衝式頻率和幅度滴注GnRH,才能使血中LH與FSH濃度呈現類似正常的脈衝式波動,從而激發青春期發育。看來,激素呈脈衝式釋放對發揮其作用是十分重要的。腺垂體的促性腺激素細胞的膜上有GnRH受體,GnRH與其受體結合后,可能是通過磷脂醯肌醇信息傳遞系統導致細胞內Ca2濃度增加而發揮作用的。在人的下丘腦,GnRH主要集中在弓狀核、內側視前區與室旁核。除下丘腦外,在腦的其他區域如間腦、邊緣葉,以及松果體、卵巢、睾丸、胎盤等組織中,也存在著GnRH。GnRH對性腺的直接作用則是抑制性的,特別是藥理劑理的GnRH,其抑制作用更為明顯,對卵巢可抑制卵泡發育和排卵,使雌激素與孕激素生成減少;對睾丸則抑制精子的生成,使睾酮的分泌減低。
(三)生長抑素與生長素釋放激素
1.生長抑素(生長素釋放抑制素,growthhormonerelease-inlease-inhibitinghormoneGHRIH或somatostatin)是由116個氨基酸的大分子肽裂解而來的十四肽,其分了結構呈環狀,在第3位和第14位半胱氨酸之間有一個二硫鍵,其化學結構為:生長抑素是作用比較廣泛的一種神經激素,它的主要作用是抑制垂體生長素(GH)的基礎分泌,也抑制腺垂體對多種刺激所引起的GH分泌反應,包括運動、進餐、應激、低血糖等。另外,生長抑素還可抑制LH、FSH、TSH、PRL及ACTH的分泌。生長抑素與腺垂體生長素細胞的膜受體結合后,通過減少細胞內cAMP和Ca2而發揮作用。除下丘腦外,其他部位如大腦皮層、紋狀體、杏仁核、海馬,以及脊髓、交感神經、胃腸、胰島、腎、甲狀腺與甲狀旁腺等組織廣泛存在生長抑素。在腦與胃腸又純化出28個氨基酸組成的在GHRIH28,它是GHRIH14N端向外延伸而成。生長抑素的垂體外作用比較複雜,它在神經系統可能起遞質或調質的作用;生長抑素對胃腸運動與消化道激素的分泌均有一定的抑制作用;它還抑制胰島素、胰高血糖素、腎素、甲狀旁腺激素以及降鈣素的分泌。2.生長素釋放激素(growthhormonereleasinghormoneGHRHA)由於下丘腦中GHRH的含量極少,致化學提取困難。1982年有人首先從一例患胰腺癌伴發肢端肥大症患者的癌組織中提取並純化出一種44個氨基酸的肽,它在整體和離體實驗均顯示有促GH分泌的生物活性。1983年,從大鼠下丘腦中提純了GHRH43,這種四十三肽對人的腺垂體也有很強有促GH分泌作用。近年用DNA重組扶得到GHRH40和GHRH44的基因,這些基因已被克隆化,並非酵母系統中傳代和表達,為提供充足與兼價的GHRH開拓了可喜的前景。產生GHRH的神經元主要分佈在下丘腦弓狀核及腹內側核,它們的軸突投射到正中隆起,終止於垂體門脈初級毛細血管旁。GHRH呈脈衝式釋放,從而導致腺垂體的GH分泌也呈現脈衝式。大鼠實驗證明,注射GHRH抗體后,可消除血中GH濃度的脈衝式波動。一般認為,GHRH是GH分泌的經常性調節者,而GHRIH則是在應激刺激GH分泌過多時,才顯著地發揮對GH分泌的抑制作用。GHRH與GHRIH相互配合,共同調節腺垂體GH的分泌。在腺垂體生長素細胞的膜上有GHRH受體,GHRH與其受體結合后,通過增加內cAMP與Ca2促進GH釋放。
(四)促腎上腺皮質激素釋放激素
促腎上腺皮質激素釋放激素(corticotropinreleasinghormone,CRH)為四十一肽,其主要作用是促進腺垂體合成與釋放促腎上腺皮質激素(ACTH)。腺垂體中存在大分子的促阿片-黑素細胞皮質素原(pro-opiomelanocortin,POMC),簡稱阿黑皮素原。在CRHA作用下經酶分解了ACTH、溶脂激素(lipotropin,β-LPH)和少量的β-內啡肽。靜脈注射CRH5-20min后,血中ACTH濃度增加5-20倍。分泌CRH的神經元主要分佈在下丘腦室旁核,其軸突多投射到正中隆起。在下丘腦以外部位,如杏仁核、海馬、中腦,以及松果體、胃腸、胰腺、腎上腺、胎盤等處組織中,均發現有CRH存在。下丘腦CRH以脈衝式釋放,並呈現晝夜周期節律,其釋放量在6-8點鐘達高峰,在0點最低。這與ACTH及皮質醇的分泌節律同步。機體遇到的應激刺激,如低血溏、失血、劇痛以及精神緊張等,作用於神經系統不同部位,最後將信息彙集於下丘腦CRH神經元,然後通過CRH引起垂體-腎上腺皮質系統反應。CRH與腺垂體促腎上腺皮質激素細胞的膜上CRH受體結合,通過增加細胞內cAMP與Ca2促進ACTH的釋放。
(五)催乳素釋放抑制因子與催乳素釋放因子
下丘腦對腺垂體催乳素(PRL)的分泌有抑制和促進兩種作用,但平時以抑制作用為主。首先在哺乳動物下丘腦提取液中,發現一種可抑制腺垂體釋放PRL的物質,稱為催乳素釋放抑制因子(prolactinrelease-inhibitingfactorPIF)。隨後,又在下丘腦提取液中發現還有一咱能促進腺垂體釋放PRL的因子,稱為催乳素釋放因子(prolactinreleasingfactorPRF)。將下丘腦提取液中的TRH分離出去,仍具有PRF活性,說明下丘腦提取液中PRF活性不是來自TRH。PIF與PRF的化學結構尚不清楚,由於多巴肽可直接抑制腺垂體PRL分泌,注射多巴胺可使正常人或高催乳素血症患者血中的PRL明顯下降,而且在下丘腦和垂體存在的多巴胺,因此有人進出多巴胺可能就是PIF的觀點。
(六)促黑素細胞激素釋放因子與抑制因
促黑素細胞激素釋放因子(melanophore-stimulatinghormonereleasingfactorMRF)(melanophore-stimulatinghormonerelease-inhibitingfactorMIF)可能是催產素裂解出來的兩種小分子肽。MRF促進MSH的釋放,而MIF則抑制MSH的釋放。
下丘腦位於丘腦下鉤的下方,構成第三腦室的下壁,界限不甚分明,向下延伸與垂體柄相連。下丘腦面積雖小,但接受很多神經衝動,故為內分泌系統和神經系統的中心。它們能調節垂體前葉功能,合成神經垂體激素及控制自主神經和植物神經功能。下丘腦的神經分泌物是通過門脈流入垂體前葉的,有的激發垂體前葉的釋放,稱釋放激素(RH);有的抑制垂體前葉激素的釋放,稱抑制激素(IH)。抑制的促激素釋放或抑制激素有:促甲狀腺激素釋放素(TRH)、促腎上腺皮質激素釋放激素(cRH)、促卵泡生成激素釋放激素(FSH-RH)、促黃體生成激素釋放激素(LH-RH)、生長激素釋放激素(GRH)、生長激素抑制激素(GIH或S.S.)、泌乳激素釋放激(PRH)、黑色細胞刺激素抑制激素(MRIH)及黑色細胞刺激素釋放激素(MRH)等十種。下丘腦分泌的釋放抑制激素、垂體分泌的促激素和靶腺合成的激素,形成一個激素網,調節著機體的許多活動。生理功能下丘腦是大腦皮層下調節內臟活動的高級中樞,它把內臟活動與其他生理活動聯繫起來,調節著體溫、攝食、水平衡、血糖和內分泌腺活動等重要的生理功能。體溫調節動物實驗中觀察到,在下丘腦以下橫切腦幹后,其體溫就不能保持相對穩定;若在間腦以上切除大腦後,體溫調節仍能維持相對穩定。現已肯定,體溫調節中樞在下丘腦;下丘腦前部是溫度敏感神經元的所在部位,它們感受著體內溫度的變化;下丘腦後部是體溫調節的整合部位,能調整機體的產熱和散熱過程,以保持體溫穩定於一定水平。攝食行為調節用埋藏電極刺激清醒動物下丘腦外側區,則引致動物多食,而破壞此區后,則動物拒食;電刺激下丘腦腹 下丘腦(垂體)內側核則動物拒食,破壞此核后,則動物食慾增大而逐漸肥胖。由此認為,下丘腦外側區存在攝食中樞,而腹內側核存在所謂飽中樞,後者可以抑制前者的活動。用微電極分別記錄下丘腦外側區和腹內側核的神經元放電,觀察到動物在飢餓情況下,前者放電頻率較高而後者放電頻率較低;靜脈注入葡萄糖后,則前者放電頻率減少而後者放電頻率增多。說明攝食中樞與飽中樞的神經元活動具有相互制約的關係,而且這些神經元對血糖敏感,血糖水平的高低可能調節著攝食中樞和飽中樞的活動。水平衡調節水平衡包括水的攝入與排出兩個方面,人體通過渴感引起攝水,而排水則主要取決於腎臟的活動。損壞下丘腦可引致煩渴與多尿,說明下丘腦對水的攝入與排出均有關係。下丘腦內控制攝水的區域與上述攝食中樞極為靠近。破壞下丘腦外側區后,動物除拒食外,飲水也明顯減少;刺激下丘腦外側區某些部位,則可引致動物飲水增多。下丘腦控制排水的功能是通過改變抗利尿激素的分泌來完成的。下丘腦內存在著滲透壓感受器,它能感受血液的晶體滲透壓變化來調節抗利尿激素的分泌;滲透壓感受器和抗利尿激素合成的神經元均在視上核和室旁核內。一般認為,下丘腦控制攝水的區域與控制抗利尿激素分泌的核團在功能上是有聯繫的,兩者協同調節著水平衡。調節腺垂體激素分泌下丘腦的神經分泌小細胞能合成調節腺垂體激素激素分泌的肽類化學物質,稱為下丘腦調節肽。這些調節肽在合成后即經軸突運輸並分泌到正中隆起,由此經垂體門脈系統到達腺垂體,促進或抑制某種腺垂體激素的分泌。下丘腦調節肽已知的有九種:促甲狀腺激素釋放激素、促性腺素釋放激素、生長素釋放抑制激素、生長素釋放激素、促腎上腺皮質激素釋放激素、促黑素細胞激素釋放因子、促黑色細胞激素釋放抑制因子,催乳素釋放因子、催乳素釋放抑制因子。對情緒反應的影響下丘腦內存在所謂防禦反應區,它主要位於下丘腦近中線兩旁的腹內側區。在動物麻醉條件下,電刺激該區可獲得骨骼肌的舒血管效應(通過交感膽鹼能舒血管纖維),同時伴有血壓上升、皮膚及小腸血管收縮、心率加速和其他交感神經性反應。在動物清醒條件下,電刺激該區還可出現防禦性行為。在人類,下丘腦的疾病也往往伴隨著不正常的情緒反應。對生物節律的控制下丘腦視交叉上核的神經元具有日周期節律活動,這個核團是體內日周期節律活動的控制中心。破壞動物的視交叉上核,原有的一些日周期節律性活動,如飲水、排尿等的日周期即喪失。視交叉上核可能通過視網膜-視交叉上核束,來感受外界環境光暗信號的變化,使機體的生物節律與環境的光暗變化同步起來;如果這條神經通路被切斷,視交叉上核的節律活動就不再能與外界環境的光暗變化發生同步。
簡介
下丘腦疾病是由於多處原因所致下丘腦功能損的一組疾病,主要特點是內分泌功能紊亂與植物神經功能失調。下丘腦既是一高級植物神經中樞,也是一功能複雜的高級內分泌中樞。下丘腦與垂體功能,性腺活動,體溫調節,食慾控制及水的代謝均有極密切的關係。下丘腦神經可為膽鹼能性,多巴胺能性或腎上腺素能性,並且在同一解剖位點上往往顯示不同神經遞質的化學染色,提示不同的生理功能在同一區域的重疊。下丘腦分泌的刺激垂體前葉的激素有促甲狀腺生長激素釋放激素(TRH)、促黃體激素釋放激素(LHRH)、促卵泡激素釋放激素(FSHRH)、生長激素釋放激素(GHRH)、促皮質激素釋放激素(CRH)、催乳素釋放因子(PRF);其抑制性的激素則有催乳素抑制因子(PIF),生長激素抑制激素(SST)。其分泌作用在垂體後葉的激素有血管加壓素(VP),抗利尿激素(ADH)。
病理
一、先天性(一)生長激素缺乏(伴有或不伴有其他激素缺乏)(二)促黃體激素(LH)和促卵泡激素(FSH)缺乏。(三)性幼稚-色素性視網膜炎-性發育不全綜合征(Laurence-Moon-BiedIe syndrome)
二、腫瘤
三、感染、腦炎、結核病、梅毒。
四、肉芽腫,結節病,漢-許-克病(Hand-Schuller-Christian disease),嗜酸細胞肉芽腫,多系統性肉芽腫。
六、機械壓迫,各種原因的腦積水。
七、醫源性,放射治療,神經外科手術。
八、創傷,顱腦外傷。
內分泌功能異常
(一)垂體激素分泌不足。多見於各種原因贊成的垂體柄損傷。當垂體柄被阻斷後,除PRL外的任何垂體激素均可缺乏或不足,常見的有尿崩症和伴有高泌乳素血症的性腺功能低下。此外,還可引起甲狀腺功能減退,腎上腺皮質功能減退。生長激素(GH)分泌減少。本組疾病相應的垂體激素的基礎測定與動力學試驗均表現為分泌低下,給於適量的釋放激素,可呈現良好的反應。
(二)垂體激素分泌過多。CRH分泌過量是柯興病(Cushing disease)的病因。GHRH分泌過多,導致肢端肥大症。較早地分泌過多的GnRH引起垂體促性腺激素(Gn)的過早釋放,可以導致真性青春期早熟。先天性囊性纖維性骨炎綜合征(AIbright syndrome)可合併性早熟。TRH分泌增多可致下丘腦性甲狀腺機能亢進症。
(三)激素節律性分泌失常。ACTH分泌的日節律可因某些下丘腦疾病和柯興綜合征的影響而消失;其他有日節律分泌的激素GH和PRL和按月節律分泌的激素LH與FSH均可因下丘腦疾病失去分泌的固有節律。
(四)青春期發育異常。下丘腦後部的疾病能消除對垂體分泌Gn的抑制作用,導致青春期性早熟。反之,下丘腦疾病也可引起青春期延緩。
下丘腦癥狀
(一)肥胖。患者由於腹正中核的飽食中樞失去功能,以致食慾增加而肥胖。肥胖可以是本症突出的和唯一的表現,過度肥胖的病人其體重往往可以持續增加,引起這種現象的原因不甚清楚。Prade-WiIIi 綜合征是由於下丘腦功能異常,有顯著的肥胖、多食、糖尿病、表情遲鈍、性腺功能低下和小手小腳。在性幼稚-色素性視網膜炎-多指畸形綜合征,肥胖也是突出的癥狀之一。
(二)厭食與消瘦。當腹外側核飲食中樞受損后,可致厭食和消瘦、嚴重者呈惡液質,肌肉無力,毛髮脫落。重症還可伴發垂體前葉功能減退。
(三)睡眠異常
①發作性睡病,最為常見,每次發作持續數分鐘至數小時不等,難以抗拒。
②深睡眠症可持續睡眠數天至數周,可叫醒進食,排尿,而後又入睡。
③發作性嗜睡貪食症,可持續睡眠數小時至數天,醒后貪食,多肥胖。
④夜間頑固性失眠。
(四)體溫調節障礙。低體溫較高體溫多見。下丘腦性低體溫程度多取決於環境的濕度的影響。患者對寒冷缺乏政黨的代償機制。高體溫對退熱葯無效。
(五)水平衡的調節障礙。視上核受損可致尿崩症。如果下丘腦的口渴中樞受累,可引起液體攝入減少,可導致脫水和血清鈉、氯的升高,可為補足液體和血管加壓素所糾正。
其他
(一)頭痛與視野缺損。多與下丘腦疾病的性質有關。下丘腦疾病所致的頭痛與視野缺損較垂體疾病為多。
(二)行為異常。下丘腦腹外側核及視前區病變時可產生行為與精神異常,患者多有行為動作減少,甚至終日靜坐不動。常伴有定向力障礙,喜怒無常,幻覺等。
基本信息
- 中文名
- 下丘腦
- 別名
- 丘腦下部
- 外文名
- hypothalamus
- 出處
- 人體解剖學名詞
- 重量
- 4g
- 分區
- 視上部,結節部,乳頭部
- 占腦比重
- 0.3%
- 所處位置
- 丘腦溝以下