細胞色素P450同工酶
細胞色素P450同工酶
人體內代謝藥物的主要酶是細胞色素P450超家族(Cytochrome P450 proteins, CYP),它們是一類主要存在於肝臟、腸道中的單加氧酶,多位於細胞內質網上,催化多種內、外源物質的(包括大多數臨床藥物)代謝。P450酶能通過其結構中的血紅素中的鐵離子傳遞電子,氧化異源物,增強異源物質的水溶性,使它們更易排出體外。 CYP有多個亞家族,包括CYP3A4,CYP3A5, CYP2D6,CYP2C9,CYP2C19等。
細胞色素P450同工酶是血紅蛋白超級家族,它是內質網膜上混合功能氧化酶系統的末端氧化酶。稱呼這些為同工酶是由於其與一氧化碳結合和還原時,分光光度法測得的吸收峰在450nm附近。P450同工酶認為已存在了350萬年,在細菌、真菌及動植物中均可發現其存在。現在已清楚認為人類細胞色素酶參與外源性物質(如藥物、酒精、抗氧化劑、有機溶劑、麻醉藥、染料、環境污染物質、化學製品)的代謝,所產生的代謝產物可能有毒性或致癌性。它們在氧化、過氧化和還原內源性生理化合物,如甾體、膽汁酸、脂肪酸、前列腺素、生物源性氨類和retinoids等代謝起重要作用。最近除了研究細胞色素同工酶參與代謝以外,還揭示了其新的生理及病理作用。
當將CO氣泡通過由聯二亞硫酸鈉所還原的鼠肝微粒體懸浮液時,在該懸浮液的差視光譜中可出現一個峰值在450nm的強吸收峰,該吸收峰與其它血紅蛋白/一氧化碳結合物的吸收峰位置不同(後者峰位在420nm左右)。這種色素有Omura和Sato表徵為細胞色素,並命名為P450,意即一種在450nm處有最大吸收峰的細胞色素,P是Pigment的縮寫。
早期研究證實細胞色素P450有多種類型,但並不知道不同物種和組織有相似的同功異構體。研究人員根據同工酶的光譜特性、電泳泳動度或其底物將其分別命名。隨著人們認識氨基酸順序的迅速進展,Nehert及其同事在常見氨基酸順序的基礎上提出了通用的系統命名法,后被廣泛接受。其將同工酶及基因分為家族和亞家族,以"CYP"為詞首來命名所有物種的細胞色素P450同工酶(除果蠅及鼠基因用'Cyp'外)。在該系統中,所有來源的細胞色素P450蛋白的氨基酸若有40%以上的同一性,則歸於同一家族,並以阿拉伯數字來標示。亞家族酶由氨基酸順序有55%以上相似的酶組成,以大寫字母標示,字母後面的阿拉伯數字錶示不同的酶,與酶相關的基因則用斜體字表示。比如,CYP2家族有幾個亞家族,諸如CYP2C、CYP2D、CYP2E。數字代表不同的酶,如CYP2D6,基因則用CYP2D6表示。不論其來源或催化活性為何,這種命名法的優點是很易識別結構一致或高度相似的細胞色素P450S。
有列表報道481個P450基因和22個假基因(截止到1995年10月)。描述了74個基因家族,在人類曾報道有14個基因家族,並曾發現人類基因組內有20個亞家族。
現已清楚細胞色素P450系統和肝臟代謝有關。CYP1,2,3家族約佔總肝P450含量的70%,並負責大多數藥物的代謝。根據它們在肝臟的表達,CYP3A約佔總肝P450的30%,CYP2約佔20%,CYP1A2佔13%,CYP2E1佔7%,CYP2A6佔4%,CYP2D6佔2%。在大量的組織內,包括小腸、胰、腦、肺、腎上腺、腎、骨髓、肥大細胞、皮膚、卵巢及睾丸均發現有其他的肝細胞色素P450。
整個腸道粘膜內,細胞色素P450同工酶存在有不同的數量。CYP3A4存在於小腸粘膜,CYP1A1存在於十二指腸。十二指腸及空腸有少量的CYP2D6及CYP2C8-10。非那西丁和氟西泮(flurasepam)系經腸道細胞色素P450酶代謝。
整個腦內均發現有細胞色素P450同工酶,且腦幹及小腦的濃度很高,認為在調節皮質酮和孕酮的濃度非常重要。皮質酮和孕酮在應激、妊娠及月經周期中性情變化、睡與醒周期中有一定的作用。
近端腎小管腔內的微絨毛狀緣及腎髓質也發現有細胞色素P450同工酶。它們可能催化腎內花生四稀酸導致產生成二十烷類物質,後者有血管活性,並可影響鐵離子的轉運,然後影響調控液體容量和成分的生理機制。肺泡II型細胞也發現細胞色素P450同工酶,但酶的總量相對較少,因此對代謝的作用不大。脂肪細胞內含有高濃度CYP19酶,認為在老年人雌激素的合成中可能起重要作用。
遺傳多態性是一種孟德爾遺傳特性,正常群體中至少存在兩種表型,每一種的頻率均不低於1%。弱或慢代謝者具有純合的常染色體隱性等位基因(常為突變的等位基因)而強或快代謝者具有雜合的或純合的顯性等位基因。一些細胞色素P450同工酶與遺傳多態性有關。最為人知的多態性同工酶是CYP2D6即異喹胍羥化酶。CYP2D6基因位於第22染色體,該基因的突變會導致酶的親和力減少或降低。5-10%的高加索人或0.9%的亞洲人的異喹胍和CYP2D6其它底物的代謝率明顯降低。除了弱和強代謝者,現在還發現一組稱為超快速代謝者。這組與活性極高的從基因放大而來的CYP2D6有關。
CYP2C19酶首先於S-美芬妥英(s-mephenytoin)的羥化中認識,也與遺傳多態性有關。S-美芬妥英慢代謝在高加索人為2-5%,日本人為20%,非洲的美洲人為19%,非洲人為8%。
CYP2E1酶也示有遺傳多態性,可影響人細胞色素基因CYP2E1 51-側面區,(含DNA的非編碼基因區,與蛋白質相互作用以促進或抑制轉錄)。它含有一些多態性長度限制性碎片,可影響轉錄或蛋白質表達的功能活性,並與酒精性肝病、吸煙引起的肝、肺癌的發展有關。
藥物代謝遺傳多態性的臨床重要性取決於藥物的活性是依賴於底物還是代謝物,以及影響藥物總的消除通路的程度。異喹胍的抗高血壓作用存在於藥物自身而其消除取決於通路。因此,正常劑量下,弱代謝個體就有更大的可能出現明顯的和長期的副作用。相反,可待因的鎮痛活性是因為被CYP2D6(異喹胍羥化酶)脫甲基后形成嗎啡所致。弱代謝者其鎮痛效能就較低。另外,涉及代謝酶抑制或誘導的藥物相互作用程度受遺傳因素的影響且可以預測。奎尼丁是異喹胍羥化酶的強效抑製劑,它可使正常有效代謝變為表型的弱代謝者,從而增加潛在的副作用,有人建議在口服治療指數窄的藥物前,對病人的特定代謝路徑進行表型研究。表型以給予單一劑量的標誌物后藥物及其代謝物在尿內濃度比率為基礎的。金雀花鹼(sparteine)、異喹胍(debrisoquine)及右美沙芬(dextromethorphan)均可作為CYP2D6多態性的標誌物。其它標誌物有CYP1A2 的咖啡因,CYP3A4的利多卡因。代謝表型的應用不廣泛,主要因為其昂貴,不方便以及特異性及敏感性均較低。用多聚酶鏈反應對外周淋巴細胞的DNA分析所進行的基因型研究可直接預測代謝表型。目前,DNA檢驗可預測出95%的健康志願者有CYP2D6表型。
肝臟的細胞色素P450系統在藥物的代謝中起著重要作用,它將藥物由疏水型轉化為更易排泄的親水型。生物轉化反應一般可分為細胞色素P450依賴的I相和II相結合反應。最近幾年內已清楚藥物的肝外代謝對體內藥物總的降解起一定的作用。胃腸道內的生物轉化是非常重要的,因為它可降低口服藥物的生物利用度。遺傳多態性、酶抑制、酶誘導及生理因素均可引起細胞色素P450活性的改變。這有一定的臨床含意,因為這會引起藥物葯代動力學的改變,導致藥物效能改變,因代謝減少或毒性代謝物生成增多而增加藥物毒性,以及導致藥物的相互作用。
CYP1A家族以兩個基因為特性,主要參與多環芳香族化合物及芳香胺的代謝。肝及肝外組織,如肺,發現有低濃度的CYP1A1(芳基族碳氫化合物羥化酶),可被多環碳氫化合物誘導。肝內誘導機制已被廣泛研究。誘導劑彌散入肝細胞,然後與載體蛋白受體結合。載體蛋白複合物進入核內與DNA結合后,促進mRNA的轉錄,後者離開核並促進粗面內質網生成細胞色素酶。一氧化氮(NO)可與細胞色素P450酶的催化型血紅素部分結合。體外研究用V79中國倉鼠細胞(可穩定表達人CYPIA1和CYP1A2酶)表明CYP1A1對NO的抑制作用較為敏感。
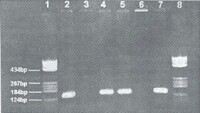
細胞色素P450基因與食管癌易感性的關係
CYP2C亞家族酶可使安定、三環類抗抑鬱葯(阿米替林、丙米嗪)去甲基,還可氧化奧美拉唑。2C9的底物是苯妥英、甲苯磺丁脲(tolbutamine)和雙氯芬酸(diclofenec)。2C10酶參與甲苯磺丁脲和環已巴比妥的羥化。苯磺唑酮(sulfinpyrazone)和硫苯拉唑(sulfaphenaxole)可抑制這兩種酶。2C19參與環已巴比妥、安定和S-美芬妥英(S-mephenytoin)的代謝。
對2D6酶(異喹胍(debrisoquine)羥化酶)的研究很多,它參與大量藥物的代謝,如異喹胍、三環類抗抑鬱葯、選擇性5-羥色胺再攝取抑製劑(除外fluvoxamine)、鎮痛葯(可待因、右美沙芬(dextromethorphan))、抗心律失常葯(金雀花鹼(sparteine)、恩卡尼encainiide、氟卡龍flecanide)、β-阻滯劑(美托洛爾metoprolol)和ondansetron。CYP2D6酶的活性呈雙峰分佈,提示人群中顯性和隱性等位基因的存在。藥物對同工酶無誘導作用。奎尼丁和選擇性5-羥色胺再攝取抑製劑可與該酶緊密結合併產生抑制作用。口服由CYP2D6代謝的藥物的病人,可被奎尼丁有效的變為弱代謝者。選擇性5-羥色胺,如paroxetine 和fluoxetine是強效競爭性、可逆性抑製劑,而sertraline和fluvoxamine的作用程度則較低。
CYP2E亞家族僅包括一個基因,是毒理學中一個重要的酶。CYP2E1(二甲基亞硝胺、N-去甲基酶),負責許多揮發性麻醉藥(如七氟醚、安氟醚、甲氧氟烷、異氟醚、乙醚、三氯乙烯和氯仿、乙醇及芳香類化合物,如苯、撲熱息痛及亞硝基二甲胺)的代謝。異煙肼和乙醇可誘導CYP2E1,並為戒酒硫(Disulfiram)所抑制,慢性乙醇消耗(因CY2E1誘導的結果)可增加苯巴比妥、甲苯磺丁脲、propanolol 和利福平的代謝率。相反,急性酗酒可競爭性的抑制2E1,從而降低苯二氮卓類、吩噻嗪類、巴比妥類、嗎啡和華法林的代謝。
CYP3家族參與大量內源性和外源性化合物的代謝,可被類固醇、大環內酯類(macrolide)抗生素、抗真菌葯和苯巴比妥所誘導。可為3A亞家族所代謝的內源性化合物包括孕酮、oestradil 、皮質醇、睾丸酮和雄稀二酮。這些酶代謝的外源性化合物包括紅黴素、利多卡因、咪噠唑侖、阿芬太尼、環孢黴素(cyclosporine)和二氫嘧啶(尼非地平)。這些底物的主要代謝途徑包括氧化、去烷基化、硝基還原和羥化。這些酶約佔肝內P450含量的30%,腸壁P450含量的70%。人的這個亞家族受CYP3A3、3A4、3A5、3A7四個基因控制。CYP3A4是肝內最常見最豐富的細胞色素亞家族。CYP3A4是人體中芬太尼氧化為非芬太尼的主要催化劑,在鼠則為3A1/2。儘管CYP1A2催化羅比卡因(ropivacaine)代謝至3-OH ropivacaine。但在體內主要代謝物(4-OH ropivacaine,2-OH ropivacaine和2-6-pipecoloxylidide ),由CYP3A4催化。CYP3A4也負責肝內將利多卡因代謝為inono ethylglycinexylide。咪噠唑侖顯著的首過氧化作用是由小腸粘膜的CYP3A4代謝產生的。CYP3A7僅發現於胎兒肝內。利福平是CYP3A酶最有力的誘導劑,其它CYP3A的誘導劑還有苯巴比妥和苯妥英。酮康唑、醋竹桃黴素、紅黴素、西咪替丁、葡萄汁可抑制CYP3A酶。關於CYP3A遺傳多態性的可能性尚未確認。
CYP3A4酶參與多種麻醉藥物的代謝:如阿片類(芬太尼、蘇芬太尼、阿芬太尼、右美沙芬)、苯二氮卓類(咪噠唑侖、安定)、和局麻藥(利多卡因、羅比卡因)。因而這些麻醉藥清除率個體間變異可能是由於CYP3A4表達的差異和CYP3A4相關藥物的相互作用。最近研究報道誘導時聯合給予芬太尼可使咪噠唑侖的清除半衰期延長50%,清除減少30%,可能是引起CYP3A活性的競爭性抑制所致。異丙酚可通過抑制CYP2B1和CYPIA1而干擾阿芬太尼和蘇芬太尼的代謝。異丙酚與酶的血紅素部分結合對CYP2B1和CYPIA1的抑制呈濃度依賴性。由於異丙酚分子太大不能與酶的活性部位有效結合,因此其抑制CYP2E1的作用是有限的。因此異丙酚可改變聯合用藥,如阿芬太尼、蘇芬太尼的代謝,但它不能有效抑制由CYP2E1參與的揮發性藥物(安氟醚、七氟醚、和甲氧氟烷)的代謝。
細胞色素P450酶可代謝揮發性氟類麻醉藥(安氟醚、七氟醚、異氟醚、和甲氧氟烷)。CYP2E1是肝內三氯乙烯、氟烷、安氟醚、七氟醚代謝的重要酶。約5%的七氟醚可很快代謝為氟化物和hexafluoro isopropanol (HFIP),再被糖醛酸化后經尿排泄。不過除了肝CYP2E1,其它P450酶如CYP1A2、2C及2D6也參與甲氧氟烷的代謝,七氟醚和甲氧氟烷經人腎微粒體脫氟生成的氟離子數量取決於時間、麻醉藥濃度和NADPH濃度。腎CYP2E1、2A6及3A對於腎代謝甲氧氟烷和七氟醚有一定關係。腎內甲氧氟烷脫氟的速率比七氟醚快3-10倍,然後生成氟離子,並增加腎內氟化物的濃度和產生潛在腎毒性,腎細胞色素同工酶代謝的七氟醚較少。使得腎內氟化物的濃度較低,血漿氟離子的濃度並不能用於預測所有鹵代類吸入麻醉藥的腎毒性。
最近的實驗表明CYP2E1主要催化氟烷生成三氟乙醯基蛋白。後者與引起氟烷肝炎的靶抗原有關。
妊娠、哺乳及酒精可使腦內細胞色素P450的濃度升高。妊娠可誘導CYP2D6並導致酶的活性增加,酒精可使CYP2C、2E1、4A的濃度增加3-5倍,這提示酒精、甾類麻醉藥及其它外源性精神藥物可能在腦內代謝。同時也提出腦細胞色素P450在對外源性藥物耐受性及神經毒性研究中的作用問題。
CYP1A2和3A4同工酶對醯胺類局麻藥於肝內的代謝非常重要。Ropicaaine 代謝時,CYP1A2負責3-OH-ropivacaine 的生成,而CYP3A負責pipecoloxglidide 的生成。CYP3A4可將利多卡因代謝成mono ethylglycinexylide。
有些口服鎮痛葯可代謝成為活性代謝物。CYP2D6可將曲馬多代謝為[(+)-M1],此代謝產物與u-阿片受體結合產生鎮痛作用。CYP2D6也對可待因和氫可酮的鎮痛作用很重要通過O-去甲基,兩者分別形成了嗎啡和氫嗎啡酮。但是,這種作用可被美沙酮抑制。更主要的是使用美沙酮治療濫用嗎啡時可引起未能料及的酶底物的藥物中毒。
過去的幾年來已明白細胞色素P450酶參與某些內源性化合物,如甾體類激素、膽固醇及脂肪酸的生物合成和/或降解。
最近的研究提示細胞色素P450酶在腦內有一定的生理作用,如經花生四稀酸代謝物的信號傳導,可能參與下丘腦及垂體釋放肽類激素;通過花生四稀酸代謝物調節腦血管張力;調節腦內的孕酮及皮質類固醇,此兩種物質被認為通過與GABA受體的相互作用影響情緒和覺醒狀態;控制細胞內膽固醇濃度,可影響低密度脂蛋白受體及參與膽固醇合成的酶的轉錄。曾提出CYP2D6酶調節神經遞質,如多巴胺及5-羥色胺的代謝和過程,因此可能在決定個體的精神狀態及個性中有一定的作用。
CYP11A、CYP11B、CYP17、CYP19和CYP21酶可影響腎上腺及性腺中類固醇的產生。腎上腺類固醇(皮質醇及醛固酮)對於維持電解質及糖的平衡很重要,而性腺類固醇(脫氫雄甾酮及雄稀二酮)則負責生殖功能和性徵差異。CYP19在性腺、腦、胎盤及脂肪組織合成雌激素起著重要作用。
細胞色素P450系統在花生四稀酸代謝中起著重要作用,在此領域已進一步加強研究。來源於細胞色素P450酶(1A1、1A2、2B1、2B2、2C11)調節路徑的代謝物,對於決定膜的離子通透性、酶的活性以及膜的更新可能起重要作用。在腎,腎細胞色素P450酶產生的花生四稀酸代謝產物可增強Na+ K+ATP酶及Na+K+2Cl-的共同轉運,從而產生利尿及利鈉作用。這些代謝物均顯示有血管收縮及舒張活性。從這些作用推論,細胞色素P450系統在整合體液容量及成分以及血壓的調節中起重要作用。鹽負荷可誘導腎CYP2C異構體,增加花生四稀酸代謝物,從而抑制近端和遠端曲小管Na+的重吸收,提示對鹽的攝取增加有適應作用。
在肝臟,CYP7、17、19、21和27參與膽汁酸及內源性類固醇的生物合成,從膽固醇合成膽汁酸可有二條路徑。一條由肝內CYP7(膽固醇7-α羥化酶)啟動,另一條則是由廣泛分佈的微粒體酶CYP27(固醇27-羥化酶)啟動,主要是在血管內皮細胞。人的CYP2C8酶似負責視黃醇及視黃酸(Retinoic acid)的代謝。
每個細胞色素P450同工酶由一個蛋白質及一個血紅素基彌補部分組成。細胞色素P450酶系統催化內源性和外源性化合物的代謝。內源性和外源性底物的生物轉化使這些化合物具有親水性或極性,然後才能被排泄。反應分為1相反應和2相反應。在1相反應中,細胞色素P450酶介導氧化或去甲基化。來源於煙醯胺腺嘌呤二核苷酸磷酸還原型的電子轉移反應后,血紅素彌補基與氧結合,這一反應給底物加入一分子氧。細胞色素P450催化的典型單氧化反應可歸納如下:
NADPH+H++O2+RH=NADP++H2O+R-OH
R表示底物,如甾體、脂肪酸或帶有烯烴、烷烴、芳香環或雜環取代基的化合物,常作為氧化作用的位點。藥物代謝性細胞色素P450S具有獨特的底物特異性,但常有重疊。細胞色素P450S的另一特點是在催化活性及調節上有大量物種內及物種間的變異性。一種藥物代謝涉及到一種以上的細胞色素P450同工酶。
細胞色素P450系統可催化很多反應,包括環氧化反應,N-去烷基化,O-去烷基化,S-氧化及脂肪族和芳香族殘基的羥化反應。氧化反應可使化合物激活或失活。和所有的酶一樣,細胞色素P450同工酶呈飽和Michaelos-Meuten動力學,其活性需要輔助因子,並可被誘導或抑制。多環碳氫化合物可誘導CYP1A酶,苯巴比酮可誘導CYP2B,糖皮質激素可誘導CYP3A,乙醇可誘導CYP2E及安妥明可誘導CYP4A。
許多機制可抑制細胞色素P450酶,如對細胞色素P450酶底物的單純競爭(奎尼丁,sulpaphenazole),化合物與血紅素結合(西米替丁),化合物的氧化產物與血紅素結合(醋竹桃黴素)以及直接的不可逆性失活(雙硫侖)。
藥物如速可眠(Secobarbitone),環磷醯胺(cydophosphamide)及氯黴素(chloramphenicol)被氧化成代謝產物,並與酶不可逆結合,有些藥物代謝產物對細胞色素P450的還原型(亞鐵)有高親和力,因而不會進一步氧化,認為紅黴素(erythromycin)抑制CYP3A即此之故,因而可引起阿芬太尼茶鹼及華法令的代謝減退。
藥物通過與細胞色素P450酶可逆性的互作用而產生相互作用。氟喹酮類抗生素(環丙沙星)可通過此機制抑制CYP3A4對咪噠唑侖的去甲基作用。西米替丁對氧化的非選擇性抑制作用是通過咪唑(imidazole)核的一個氮原子與細胞色素P450血紅素鐵的直接相互作用實現的。被西米替丁影響消除的藥物包括華法令、安定、苯妥英和propanolol。其他含咪唑基的藥物如酮康唑對細胞色素P450酶可能有潛在的抑制作用。雷尼替丁含有呋喃環而不是咪唑環,對細胞色素P450酶無抑制作用。
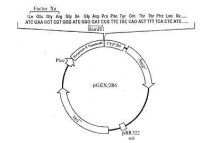
人細胞色素 P450 融合蛋白及其抗體的製備
疾病狀態中的作用
某些同工酶活性的改變對於癌的形成,腎上腺增生和帕金森氏病有一定意義。吸煙者可增加CYP1A1(芳香族碳氫化合物羥化酶)的活性,更易致發生肺癌。曾被證實個體CYP1A1的遺傳差異以致肺癌的危險性增加,含有特異性稀有等位基因純合子的個體發生疾病的危險性較高。
乳腺癌病人其CYP1B1同工酶增加。因為它參與乳房內雌激素的代謝,提示在雌激素依賴性乳房腫瘤的病因中CYP1B1起一定的作用。
在吸煙者,CYP2D6酶參與激活前致癌物,從而介導致癌作用引起肺癌的發生。CYP2E1可催化苯胺(aniline)氯化碳氫化合物和苯的代謝,引起前致癌物的形成。現提出CYP2E1的遺傳多態性對肝癌的發展可能起著重要作用。
神經元細胞內催化反應生成反應性的代謝物可能引起神經毒性導致帕金森氏病。流行病學研究指出異喹呱弱代謝者易較早發生帕金森氏病。CYP21A2遺傳編碼缺陷,可致21-羥化酶缺乏,約佔先天性腎上腺皮質增生患者的90-95%。
儘管最近對於人細胞色素P450酶的認識有很大進步,但仍需一種可靠方法來監測人類細胞色素P450的表達。使預測藥物的相互作用和藥物的代謝清除成為可能。病人所用藥物的治療指數窄時,可根據表型或基因型來指導用藥劑量和減少毒性。個別人的細胞色素P450同工酶呈特殊形式的高表達時,可以篩選確定是否易於發生癌症或毒性。因此,研究人類細胞色素P450對於藥物的治療作用和預防多種疾病(如癌)有一定的效用。

基本信息
- 中文名
- 細胞色素P450同工酶
- 外文名
- Cytochrome P450 proteins, CYP
- 存在部位
- 肝臟、腸道
- 主要酶
- 細胞色素P450超家族
- 包括
- CYP3A4,CYP3A5, CYP2D6,CYP2C9