顯生宙
指“看得見生物的年代”
顯生宙,指“看得見生物的年代”,是開始出現大量較高等動物以來的階段,包括古生代、中生代和新生代,從距今大約5.4億年前延續至今。5.4億年前,寒武紀始,生物逐漸演化出較高級的動物,動物已具有外殼和清晰的骨骼結構,故稱顯生宙。
顯生宙是從距今5.7億年至今的一段地質時期。以大量動物出現為標誌。它分為古生代、中生代和新生代。古生代包括寒武紀、奧陶紀、志留紀、泥盆紀、石炭紀和二疊紀,並可以分為早古生代及晚古生代兩部分,各包括三個紀。中生代分為三疊紀、侏羅紀、白堊紀。新生代包括第三紀和第四紀,通常把第三紀分為老第三紀和新第三紀。每個紀下可分為若干世,一般分為早、中、晚三個世或早、晚兩個世。世下再分為期。紀和世、期的劃分以地殼變動和生物群的面貌為標誌,並參以同位素年齡。
地質年代
| 時間 | 宙 | 代 | 紀 | 世 | 期 | 年代,百萬年前 |
|---|---|---|---|---|---|---|
| 顯生宙 | 新生代 | 第四紀 | 全新世 | (時:亞大西洋期、亞北方期、大西洋期、北方期、前北方期) | 0.011700 | |
| 更新世 | 晚更新期 | 0.126 | ||||
| 中更新期 | 0.781 | |||||
| 卡拉布里亞階 | 1.806 | |||||
| 格拉斯期 | 2.588 | |||||
第三紀/ 新近紀 | 上新世 | 皮亞琴察期/勃朗期 | 3.600 | |||
| 贊克爾期 | 5.333 | |||||
| 中新世 | 墨西拿期 | 7.246 | ||||
| 托爾頓期 | 11.63 | |||||
| 塞拉瓦爾期 | 13.82 | |||||
| 蘭蓋期 | 15.97 | |||||
| 布爾迪加爾期 | 20.44 | |||||
| 阿基坦期 | 23.03 | |||||
第三紀/ 古近紀 | 漸新世 | 恰特期 | 27.82 | |||
| 魯培爾期 | 33.9 | |||||
| 始新世 | 普里阿邦期 | 37.8 | ||||
| 巴爾頓期 | 41.2 | |||||
| 盧台特期 | 47.8 | |||||
| 伊普雷斯期 | 56.0 | |||||
| 古新世 | 贊尼特期 | 59.2 | ||||
| 塞蘭特期 | 61.6 | |||||
| 達寧期 | 66.0 | |||||
| 中生代 | 白堊紀 | 晚白堊世 | 馬斯特里赫特期 | 72.1 ± 0.2 | ||
| 坎帕期 | 83.6 ± 0.2 | |||||
| 桑托期 | 86.3 ± 0.5 | |||||
| 科尼亞剋期 | 89.8 ± 0.3 | |||||
| 土侖期 | 93.5 ± 0.8 | |||||
| 森諾曼期 | 99.6 ± 0.9 | |||||
| 早白堊世 | 阿爾布期 | 112.0 ± 1.0 | ||||
| 阿普第期 | 125.0 ± 1.0 | |||||
| 巴列姆期 | 130.0 ± 1.5 | |||||
| 豪特里維期 | 136.4 ± 2.0 | |||||
| 凡藍今期 | 140.2 ± 3.0 | |||||
| 貝里亞期 | 145.5 ± 4.0 | |||||
| 侏羅紀 | 晚侏羅世 | 提通期 | 150.8 ± 4.0 | |||
| 啟莫里期 | 155.7 ± 4.0 | |||||
| 牛津期 | 161.2 ± 4.0 | |||||
| 中侏羅世 | 卡洛維期 | 164.7 ± 4.0 | ||||
| 巴通期 | 167.7 ± 3.5 | |||||
| 巴柔期 | 171.6 ± 3.0 | |||||
| 阿連期 | 175.6 ± 2.0 | |||||
| 早侏羅世 | 托阿爾期 | 183.0 ± 1.5 | ||||
| 普連斯巴奇期 | 189.6 ± 1.5 | |||||
| 錫內穆期 | 196.5 ± 1.0 | |||||
| 海塔其期 | 199.6 ± 0.6 | |||||
| 三疊紀 | 晚三疊世 | 瑞替期 | 203.6 ± 1.5 | |||
| 諾利期 | 216.5 ± 2.0 | |||||
| 卡尼期 | 228.0 ± 2.0 | |||||
| 中三疊世 | 拉丁尼期 | 237.0 ± 2.0 | ||||
| 安尼西期 | 245.0 ± 1.5 | |||||
| 早三疊世 | 奧倫尼剋期 | 249.7 ± 1.5 | ||||
| 印度期 | 251.902 ± 0.024 | |||||
| 古生代 | 二疊紀 | 樂平世 | 長興期 | 254.14 ± 0.07 | ||
| 吳家坪期 | 259.1 ± 0.5 | |||||
| 瓜德魯普世 | 卡匹敦階 | 265.1 ± 0.4 | ||||
| 沃德期/卡贊期 | 268.8 ± 0.5 | |||||
| 羅德期/烏非姆期 | 272.95 ± 0.11 | |||||
| 烏拉爾世 | 空谷爾期 | 283.5 ± 0.6 | ||||
| 阿爾丁斯剋期 | 290.1 ± 0.26 | |||||
| 薩克馬爾期 | 295.0 ± 0.18 | |||||
| 阿瑟爾期 | 298.9 ± 0.15 | |||||
石炭紀/ 賓夕法尼亞紀 | 晚賓夕法尼亞世 | 格熱爾期 | 303.7 ± 0.1 | |||
| 卡西莫夫期 | 307.0 ± 0.1 | |||||
| 中賓夕法尼亞世 | 莫斯科期 | 315.2 ± 0.2 | ||||
| 早賓夕法尼亞世 | 巴什基爾期 | 323.2 ± 0.4 | ||||
石炭紀/ 密西西比紀 | 晚密西西比世 | 謝爾普霍夫期 | 330.9 ± 0.2 | |||
| 中密西西比世 | 維憲期 | 346.7 ± 0.4 | ||||
| 早密西西比世 | 圖爾奈期 | 358.9 ± 0.4 | ||||
| 泥盆紀 | 晚泥盆世 | 法門期 | 372.2 ± 1.6 | |||
| 弗拉斯期 | 382.7 ± 1.6 | |||||
| 中泥盆世 | 吉維特期 | 387.7 ± 0.8 | ||||
| 艾菲爾期 | 393.3 ± 1.2 | |||||
| 早泥盆世 | 埃姆斯期 | 407.6 ± 2.6 | ||||
| 布拉格期 | 410.8 ± 2.8 | |||||
| 洛赫科夫期 | 419.2 ± 3.2 | |||||
| 志留紀 | 普里道利世 | 無生物劃分階 | 423.0 ± 2.3 | |||
| 蘭多維列世/卡尤加世 | 盧德福德期 | 425.6 ± 0.9 | ||||
| 戈斯特期 | 427.4 ± 0.5 | |||||
| 文洛克世 | 侯默期/洛克波特期 | 430.5 ± 0.7 | ||||
| 申伍德期/托納旺達期 | 433.4 ± 0.8 | |||||
蘭多維利世/ 亞歷山大世 | 特列奇期/安大略期 | 438.5 ± 1.1 | ||||
| 愛隆期 | 440.8 ± 1.2 | |||||
| 魯丹期 | 443.4 ± 1.5 | |||||
| 奧陶紀 | 晚奧陶世 | 赫南特期 | 445.2 ± 1.4 | |||
| 凱迪期 | 453.0 ± 0.7 | |||||
| 桑比期 | 458.4 ± 0.9 | |||||
| 中奧陶世 | 達瑞威爾期 | 467.3 ± 1.1 | ||||
| 大坪期 | 470.0 ± 1.4 | |||||
| 早奧陶世 | 弗洛期 | 477.7 ± 1.4 | ||||
| 特馬豆剋期 | 485.4 ± 1.9 | |||||
| 寒武紀 | 芙蓉世 | 第十期 | c. 489.5 | |||
| 江山期 | c. 494 | |||||
| 排碧期 | c. 497 | |||||
| 第三世 | 古丈期 | c. 500.5 | ||||
| 鼓山期 | c. 504.5 | |||||
| 第五期 | c. 509 | |||||
| 第二世 | 第四期 | c. 514 | ||||
| 第三期 | c. 521 | |||||
| 紐芬蘭世 | 第二期 | c. 529 | ||||
| 幸運期 | 541.0 ± 1.0 | |||||
前寒武紀/ 隱生宙 | 元古宙 | 新元古代 | 埃迪卡拉紀 | 630 +5/-30 | ||
| 成冰紀 | 720 | |||||
| 拉伸紀 | 1000 | |||||
| 中元古代 | 狹帶紀 | 1200 | ||||
| 延展紀 | 1400 | |||||
| 蓋層紀 | 1600 | |||||
| 古元古代 | 固結紀 | 1800 | ||||
| 造山紀 | 2050 | |||||
| 層侵紀 | 2300 | |||||
| 成鐵紀 | 2500 | |||||
| 太古宙 | 新太古代 | 2800 | ||||
| 中太古代 | 3200 | |||||
| 古太古代 | 3600 | |||||
| 始太古代 | 3800 | |||||
| 冥古宙 | 雨海代 | c.3850 | ||||
| 酒神代 | c.3920 | |||||
| 原生代 | c.4150 | |||||
| 隱生代 | c.4600 | |||||
顯生宙,指“看得見生物的年代”。
自五億七千萬年前,寒武紀始,生物逐漸向較高級的發展階段進化,動物已具有外殼和清晰的骨骼結構,故稱顯生宙。顯生宙時期,就是從五億七千萬年直到現在。
顯生宙以來發生的生物進化事件主要是以多細胞動、植物出現為開始;在顯生宙的生物進化舞台上的主角,已經不再是單細胞的原核生物(主要指細菌和藻類植物)的天下,在顯生宙的初期(古生代)就出現單細胞的真核生物。因此人們又把多細胞動、植物等真核生物出現以後的地質時代稱為顯生宙。
真核生物(eukaryotes):是所有單細胞或多細胞的、其細胞具有細胞核的生物的總稱。包括所有多細胞生物——原生生物、動物、植物、真菌,以及一些單細胞的原生生物。真核生物的另一個特點是它們的細胞在製造蛋白質時可以用同一段基因製造不同的蛋白質。這個功能在術語中被稱為選擇性剪接。所有的真核生物在生物分類學上,又可稱為真核生物域,是生物分類的三域系統中之一,包括真菌界、植物界和動物界。
三域系統:三域系統是由Carl Woese(卡爾·烏斯)在1990年提出的生物分類,將原核生物分成了兩大類,起初稱為真細菌和古細菌。Woese依據16S rRNA序列上的差別,認為這兩組生物和真核生物從一個具有原始遺傳機制的共同祖先分別進化而來,因此將三者各劃為一類,作為比界高的分類系統,稱作“域”(Domain)或者“總界”(Superkingdom)。真核域。
古菌域(Archaea):古菌在生物分類學上分的一個總稱。古細菌(古菌)是原核生物中的一大類。它們既與細菌(真細菌)有很多相似之處,同時另一些特徵相似於真核生物。從rRNA進化樹上,古菌分為兩類,泉古菌(Crenarchaeota)和廣古菌(Euryarchaeota)。另外未確定的兩類分別由某些環境樣品和2002年由Karl Stetter發現的奇特的物種納古菌(Nanoarchaeum equitans)構成。Woese認為細菌、古菌和真核生物各代表了一支具有簡單遺傳機制的遠祖生物的後代。這個假說反映在了“古菌”的名稱中(希臘語archae為“古代的”)。隨後他正式稱這三支為三個域,各由幾個界組成。這種分類後來非常流行,但遠祖生物這種思想本身並未被普遍接受。一些生物學家認為古菌和真核生物產生於特化的細菌。然而﹐RNA樹有可能將進化太快的類群放錯位置(比如微孢子蟲)。有人認為生物的根應該在真細菌之內﹐很多真細菌的類群在古細菌之前分支出來﹐很晚古細菌和真核生物才彼此分開。
古菌和真核生物的關係仍然是個重要問題。除掉上面所提到的相似性,很多其他遺傳樹也將二者並在一起。在一些樹中真核生物離廣古菌比離泉古菌更近,但生物膜化學的結論相反。然而,在一些細菌,(如棲熱袍菌)中發現了和古菌類似的基因,使這些關係變得複雜起來。一些人認為真核生物起源於一個古菌和細菌的融合,二者分別成為細胞核和細胞質。這解釋了很多基因上的相似性,但在解釋細胞結構上存在困難。目前有22個古菌基因組已經完全結束了測序,另外15個的測序工作正在進行中。
真核域(Eukarya):又稱,真核生物域(或稱真核生物總界),是真核生物所屬的一個生物總稱。
真核域:動物界(Animalia)、真菌界(Fungi)、變形蟲界(Amoebozoa)、植物界(Plantae)、有孔蟲界(Foraminifera community),這是能夠在百度上到2009-12-30為至搜索到的最新分類,也有一部分人把真核生物域分為四界:原生生物界、真菌界、植物界、動物界)。由於真核生物與古菌在一些生化性質和基因相關性上具有一定相似性,因此有時也將這兩者共同歸於Neomura演化支(總域: Neomura 域)。
隨著研究的進展,分類層次不斷增加,單元上下可以附加次生單元,如總綱(超綱)、亞綱、次綱、總目(超目)、亞目、次目、總科(超科)、亞科等等。此外,還可增設新的單元,如股、群、族、組等等,其中最常設的是族,介於亞科和屬之間。
種是最小的生物單位。生物的相同科目越多,共同點也越多。
分類系統是階元系統,通常包括七個主要級別:種、屬、科、目、綱、門、界。種(物種)是基本單元,近緣的種歸合為屬,近緣的屬歸合為科,科隸於目,目隸於綱,綱隸於門,門隸於界。
例如,現代人類中的一個黃色人種,就是可以分為:真核生物域—→動物界—→脊索動物亞門—→脊椎動物總綱(超綱)—→哺乳綱—→獸亞綱—→真獸次亞綱—→靈長總目(超目)—→真靈長半目—→直鼻猴亞目—→人猿次目—→狹鼻下目—→真狹鼻小目—→人超科(總科)—→人科—→人亞科—→人族—→人屬—→人亞屬—→智人種—→現代人種—→黃色人種(蒙丁利亞人種)。
1969年美國學者魏泰克(Whitaker)也曾提出過五界分類法:原核生物界、原生生物界、真菌界、植物界、動物界。
原生生物界(Protista)是真核生物域中的一界。真核原生生物界的生物都是有細胞核的,且幾乎是單細胞生物。某些真核原生生物像植物:如矽藻(diatom);某些像動物:如變形蟲(amoeba)、纖毛蟲(ciliate);某些既像植物又像動物,如:眼蟲(euglena)。
原生生物,包括藻類(沒有歸類為原核生物域中的生物)和原生動物等。他們是由原核生物進化來的,是真核生物中最原始的類群。早期藻類是植物的祖先,早期的原生動物是動物的祖先,所以人們對生物進行分類時,常把藻類歸於植物界,把原生動物歸於動物界。在原生生物中,藻類的數量多,分佈廣,與人類的關係也最為密切。原生動物是像動物一樣的原生生物,它們會四處移動尋找食物。有些原生動物會利用長長的鞭毛,或以改變身體形狀的方式來移動
真核細胞與原核細胞的主要區別是:1真核細胞具有由染色體、核仁、核液、雙層核膜等構成的細胞核;原核細胞無核膜、核仁,故無真正的細胞核,僅有由核酸集中組成的擬核。2真核細胞的轉錄在細胞核中進行,蛋白質的合成在細胞質中進行,而原核細胞的轉錄與蛋白質的合成交聯在一起進行。3真核細胞有內質網、高爾基體、溶酶體、液泡等細胞器,原核細胞沒有。4真核生物中除某些低等類群(如甲藻等)的細胞以外,染色體上都有5種或4種組蛋白與DNA結合,形成核小體;而在原核生物則無。5真核細胞在細胞周期中有專門的DNA複製期(S期);原核細胞則沒有,其DNA複製常是連續進行的。6真核細胞的有絲分裂是原核細胞所沒有的。7真核細胞有發達的微管系統,其鞭毛(纖毛)、中心粒、紡錘體等都與微管有關,原核生物則否。8真核細胞有由肌動、肌球蛋白等構成的微纖維繫統,後者與胞質環流、吞噬作用等密切相關;而原核生物卻沒有這種系統,因而也沒有胞質環流和吞噬作用。9真核細胞的核糖體為80S型,原核生物的為70S型,兩者在化學組成和形態結構上都有明顯的區別。10真核細胞含有的線粒體,為雙層被膜所包裹,有自己特有的基因組、核酸合成系統與蛋白質合成系統,其內膜上有與氧化磷酸化相關的電子傳遞鏈。11真核生物細胞較大,一般10~100納米,原核生物細胞較小,大約1~10納米。12真核生物一般含有細胞器(線粒體和葉綠體等),原核生物的細胞器沒有膜包裹。13真核生物新陳代謝為需氧代謝(除了amitochondriats),原核生物新陳代謝類型多種多樣。14真核生物細胞壁由纖維素或幾丁質組成,動物沒有細胞壁,原核生物真細菌中為肽聚糖。15真核生物動植物中為有性的減數分裂式的受精、有絲分裂,原核生物通過一分為二或出芽生殖、裂變。16真核生物遺傳重組為減數分裂過程中的重組,原核生物為單向的基因傳遞。17真核生物鞭毛為捲曲式,主要由微管蛋白組成,原核生物鞭毛為旋轉式,由鞭毛蛋白組成。18真核生物通過線粒體進行呼吸作用,原核生物通過膜進行呼吸作用。
原核細胞功能上與線粒體相當的結構是質膜和由質膜內褶形成的結構,但後者既沒有自己特有的基因組,也沒有自己特有的合成系統。真核生物的植物含有葉綠體,它們亦為雙層膜所包裹,也有自己特有的基因組和合成系統。與光合磷酸化相關的電子傳遞系統位於由葉綠體的內膜內褶形成的片層上。原核生物中的藍細菌和光合細菌,雖然也具有進行光合作用的膜結構,稱之為類囊體,散佈於細胞質中,未被雙層膜包裹,不形成葉綠體。
最原始的真核生物的直接祖先很可能是一種異常巨大的原核生物,體內具有由質膜內褶而成的象內質網那樣的內膜系統和原始的微纖維繫統,能夠作變形運動和吞噬。以後內膜系統的一部分包圍了染色質,於是就形成了最原始的細胞核。內膜系統的其他部分則分別發展為高爾基體、溶酶體等細胞器。按照美國學者L.馬古利斯等重新提出的“內共生說”(見細胞起源),線粒體起源於胞內共生的能進行氧化磷酸化的真細菌,而葉綠體則起源於胞內共生的能進行光合作用的藍細菌。
地球上的生命,從最原始的無細胞結構生物進化為有細胞結構的原核生物,從原核生物進化為真核單細胞生物,然後按照不同方向發展,出現了真菌界、植物界和動物界。植物界從藻類到裸蕨植物再到蕨類植物、裸子植物,最後出現了被子植物。動物界從原始鞭毛蟲到多細胞動物,從原始多細胞動物到出現脊索動物,進而演化出高等脊索動物──脊椎動物。脊椎動物中的魚類又演化到兩棲類再到爬行類,從中分化出哺乳類和鳥類,哺乳類中的一支進一步發展為高等智慧生物,這就是人。
顯生宙分為古生代、中生代和新生代,其下又細分為多個紀,有寒武紀、奧陶紀、志留紀、泥盆紀、石炭紀、二疊紀、三疊紀、侏羅紀、白堊紀、第三紀、第四紀,十一個紀。
從距今約570百萬年到250百萬年,持續了320百萬年(三億二千萬年)的時間分為了,顯生宙——古生代。在古生代,動物群以海生無脊椎動物中的三葉蟲、軟體動物和棘皮動物最繁盛。在奧陶紀、志留紀、泥盆紀、石炭紀,相繼出現低等魚類、古兩棲類和古爬行類動物。魚類在泥盆紀達於全盛。石炭紀和二疊紀昆蟲和兩棲類繁盛。古植物在古生代早期以海生藻類為主,至志留紀末期,原始植物開始登上陸地。泥盆紀以裸蕨植物為主。石炭紀和二疊紀時,蕨類植物特別繁盛,形成茂密的森林,是重要的成煤期。
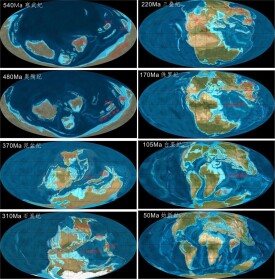
古生代的地殼運動和氣候變化深刻影響自然環境的發展。早古生代的地殼運動在歐洲稱加里東運動,在美洲稱太康運動,在中國又稱廣西運動。此時古北美、古歐洲、古亞洲、岡瓦納古陸及古太平洋、古地中海都已形成。晚古生代地殼運動在歐洲稱海西(華力西)運動,在北美稱阿勒蓋尼運動,在中國又稱天山運動。
經過古生代地殼運動,世界許多巨大的褶皺山系出現,南方的岡瓦納古陸和北方的勞亞古陸聯合在一起,形成泛古陸(聯合古陸:據推測,這一超級大陸約佔地球表面積的一半。環繞聯合古陸的原始大洋稱泛大洋或聯合古洋)。晚古生代在岡瓦納古陸發生了大規模的冰川作用,大冰蓋分佈於古南緯60°以內的今南非、阿根廷等地,該冰川作用期即地質歷史上的石炭——二疊紀大冰期。古生代的地層總稱,顯生宇——古生界。
在寒武紀時,具有硬殼的動物第一次大量地出現,許多大陸都被淺海所泛濫,超大陸剛瓦那(Gondwana)則正在南極附近形成。寒武紀晚期,地球海陸分布圖形成於前寒武世末期(大約距今六億年前)的超大陸潘諾西亞(Pannotia),在古生代的時候開始四分五裂,一個新的海洋--巨神海(Iapetus Ocean)在勞倫西亞(Laurentia,北美)、波羅地(Baltica,北歐)和西伯利亞(Siberia)這幾個古大陸之間擴張。超大陸岡瓦那(Gondwana)則在泛非褶皺帶上組合而成當時最大的大陸,範圍從赤道延伸到南極。
在奧陶世時期,溫暖的海水把石灰岩和鹽岩沉澱在岡瓦那大陸的赤道地區(Australia澳大利亞、India印度、China中國與Antarctica南極洲),同時在岡瓦那大陸的南極地區(Africa非洲與South America南美)則沉澱了冰河的沉積及冰漂的碎屑。
古生代包括了寒武紀、奧陶紀、志留紀、泥盆紀、石炭紀、二疊紀,六紀。又小分為早古生代和晚古生代,早古生代含:寒武紀、奧陶紀、志留紀;泥盆紀、石炭紀、二疊紀,又合稱晚古生代。
早古生代是海生無脊椎動物的發展時代。在早古生代海洋里生活著門類眾多的生物。植物界以海藻為主。動物界出現了三葉蟲和珊瑚、腕足類等。三葉蟲是一種節肢動物,寒武紀是三葉蟲的、全盛時代。到奧陶紀時出現了軟體動物門的頭足綱,主要生物門類還有筆石、腕足類、三葉蟲等。最值得注意的是在志留紀中期出現了脊椎動物——魚類和最早的陸生植物。例如:寒武紀的節肢動物三葉蟲、奧陶紀的筆石和頭足類、泥盆紀的珊瑚類和腕足類等。最早的脊椎動物無顎魚也在奧陶紀出現。植物以水生菌藻類為主,志留紀末期出現裸蕨植物。
鄭州、西安、開封、廣州等歷史悠久的文化古城,在考古足底下的地層時,往往會發現從上到下依次有清、明、元、宋、唐的存積或遺跡。從遠古至中古、近代到現代,歷史一層蓋過一層。
地球也很類似,不同的地層反映不同的地質世紀。只不過因為無數次山翻地復、滄海桑田,未必有清晰的層次而已。地質歷史分宙(或代)、紀、世、期。
相應地層為界、系、統、層。
冥古宙+ 太古宙+ 元古宙持續約40億年。顯生宙持續約5·7億年。不同資料年代有出入,甚至有數億年之差,這是因為最古老的生命痕迹還有許多不確定因素。
顯生宙的上限一般定在生命大爆發開始,為5·7億到6億年前。
顯生宙含:古生代、中生代、新生代。
古生代——距今約5·7億年起到約2·5億年前,持續約3·2億年
早古生代含:寒武紀、奧陶紀、志留紀。
晚古生代含:泥盆紀、石炭紀、二疊紀。
奧陶紀5·1億——4·38億年。1869英地質學家根據古代居住的民族名稱命名。
志留紀4·38億——4·1億年。名稱來源於另一個威爾士古代當地民族。
泥盆紀4·1億——3·55億年。1839將一套海成岩石層按地名命名,中文翻譯為“泥盆”。
石炭紀3·55億——2·9億年。1822研究英國地質發現穩定含煤炭地層而得名。
中生代——距今2·5億年起到6500萬年前。持續約1·85億年
中生代含:三疊紀、侏羅紀、白堊紀。
三疊紀2·5億——2·05億年。1834命名於德國西南部,那裡有三套截然不同的地層。
新生代——約距今6500萬年起到如今。新生代含:第三紀、第四紀。
早期研究地史分原始紀、第二紀和第三紀。“第三紀”由義大利地質學家1779創用。原來也將新生代稱為第三紀,1829研究法國地質時從第三紀中劃出第四紀。
第三紀6500萬——160萬年前。第三紀含:古新世、始新世、漸新世、中新世、上新世。
更新世已是人類舊石器時代了,又可細分:早更新世、中更新世、晚更新世。
全新世距今小於1·2萬年,是地質史新生代7個“新世”劃分的“滿崽”了,此時中石器時代、新石器時代相繼開始,人類使世界進入全新的時代。
新生代是地球歷史上最新的一個地質時代,它從6400萬年前開始一直持續到今天。隨著恐龍的滅絕,中生代結束,新生代開始。新生代一般被分為三個紀:古近紀、新近紀和有爭議的第四紀。這三個紀又可劃分為七個世:古新世、始新世、漸新世(屬古近紀),中新世、上新世(屬新近紀),更新世、全新世(屬第四紀)。
在過去,古近紀和新近紀常合併為第三紀,它們因而也曾分別被叫做早第三紀和晚第三紀。新生代是哺乳動物的時代。在新生代中,哺乳動物從微小簡單的原始哺乳動物發展到佔據各個生態圈的巨大的動物群。在新生代內,鳥和被子植物也有很大的發展。新生代中,盤古大陸徹底分裂,地球上的各個大陸逐漸移動到今天的位置上。
第四紀是地質時代中的最新的一個紀,它包括全新世和更新世兩個世。第四紀前是新近紀。它從約180萬年或160萬年前開始一直延續到今天。
第四紀這個名稱是1829年由儒勒·迪斯努瓦耶(Jules Desnoyers)提出的。他在研究塞納河低地的沉積層時發現了一層比新近紀更新的岩層。這個岩層一直延伸到今天。第四紀的時期基本上與最近的冰川期(包括現在的冰川回退期)相符。另一種分法是將300萬年前北極結冰的開始作為第四紀的開始,這樣的話上新世的最新的一部分也算作第四紀了。也有人不承認第四紀的存在,而將它看作第三紀的一部分。
第四紀的180或160萬年中人類的存在已被確證。在這段時間裡板塊運動小於100千米,因此可以被忽略。在這段時間裡氣候不斷變化,冰川期與冰川間期交換。在冰川期中冰川可以一直延伸到緯度40度的地方。在這段時間裡只有很少新的動物種類產生(可能因為這段時間還比較短),在更新世末期,在北半球有不少哺乳動物(如劍齒虎、猛獁象、乳齒象、雕齒獸等)滅絕。馬、駱駝等在北美洲滅絕。
中國地理學家竺可楨指出,第四紀歐洲和北美洲北部經歷了四個冰川時期和四個間冰川時期:第一冰川時期距今30萬年至27萬年;第二冰川時期距今20萬年至18萬年;第三冰川時期距今13萬年至10萬年;第四冰川時期距今6萬5千年至1萬5千年。
中生代(Mesozoic)是顯生宙的三個地質時代之一,可分為三迭紀,侏羅紀和白堊紀三個紀。中生代最早是由義大利地質學家Giovanni Arduino所建立,當時名為第二紀(Secondary),以相對於現代的第三紀。在希臘文中,中生代意為“中間的”+“生物”。中生代介於古生代與新生代之間。由於這段時期的優勢動物是爬行動物,尤其是恐龍,因此又稱為爬行動物時代(Age of the Reptiles)。
中生代也是板塊、氣候、生物演化改變極大的時代。在中生代開始時,各大陸連接為一塊超大陸-盤古大陸。盤古大陸後來分裂成南北兩片,北部大陸進一步分為北美和歐亞大陸,南部大陸分裂為南美、非洲、印度與馬達加斯加、澳大利亞和南極洲,只有澳大利亞沒有和南極洲完全分裂。中生代的氣候非常溫暖,對動物的演化產生影響。在中生代末期,已見現代生物的雛形。
古生代(Paleozoic,符號PZ)是地質時代中的一個代,開始於同位素年齡542±0.3百萬年(Ma),結束於251±0.4Ma。
古生代是顯生宙的第一個代,上一個代是元古宙的新元古代,下一個代是中生代。古生代包括了寒武紀、奧陶紀、志留紀、泥盆紀、石炭紀、二迭紀。其中寒武紀、奧陶紀、志留紀又合稱早古生代,泥盆紀、石炭紀、二迭紀又合稱晚古生代。
植物
石炭紀,尤其是上石炭紀可以被稱為是蕨類植物的時代。由於幾個立方米的木頭才能演變為一立方米的煤,因而上石炭紀時的蕨類植物森林的規模就可以從今天石煤層的規模中換算出來。這些成為今天的石煤的植物中最主要的是鱗木目和封印木屬的植物。這些樹狀的植物屬於今天的石松綱,它們可以達到40米高,其莖的直徑可達1米。屬於問荊的蘆木也達到了20米高。舌羊齒類植物也成為樹一般的高大木質植物。石炭紀末期有花植物(裸子植物如歧杉和瓦契杉)出現。
動物
海生動物:泥盆紀海中占支配作用的帶有硬骨裝甲的魚在泥盆紀-石炭紀的大滅絕後沒有再恢復過來。石炭紀海中的主要魚類是活動靈便的輻鰭魚類。
顯生宙
海百合是石炭紀出現的新生物,它們屬於棘皮動物。這些動物在石炭紀的海底形成草地一般的大面積覆蓋面,它們留下了許多化石。其它留下許多化石的動物有苔蘚蟲動物門的動物和希瓦格蜓和紡錘蜒,后兩者是單細胞動物,但它們可以達到10厘米大。
陸地動物:最早的無翼的昆蟲在下泥盆紀時代就出現了,到上石炭紀已有有翼的昆蟲。這些昆蟲還無法摺疊它們的翅膀(如蜻蜓等),石炭統煤系地層中發現超過500種的昆蟲。石炭紀時在陸地上生活的唯一的脊椎動物是兩棲動物,但它們還保存著相當的水生習性。由於它們在陸地上還沒有競爭對手,因此它們的種類非常多,有些一直大到6米長。
在上石炭紀的末期還找到了最古老的可以算作爬行動物的骨骼化石。這時也出現了最早的帶有硬殼的蛋。
是地球歷史中的一個地質時代。早在1822年石炭紀在英國就已經被看作是一個地質時代中的紀了。石炭紀的名字來自於上石炭紀時期在全世界各地形成的煤。它從3.55億年前開始,延續到2.9億年前。它與二疊紀和泥盆紀之間的邊界的年代主要是通過放射性同位素斷代獲得的。
對石炭紀的內部分類各個地區使用非常不同的系統,這些不同的系統在其使用地區和傳統中卻相當穩定。在西歐石炭紀一般被分為上下兩個亞紀,在美國分密西西比亞紀和賓西維尼亞亞紀,在俄羅斯分上中下三個亞紀。古生物學上的細節劃分一般使用海生動物:頭足綱動物、腕足綱動物、珊瑚等。陸地植物也被用來對上石炭紀作細節劃分。
古地理
出發點:在泥盆紀中北美地塊和北歐-俄羅斯地塊結合到一起。這塊大陸與後來的岡瓦那大陸的其它部分(今天的非洲、南美洲、南極洲、澳大利亞和印度)之間部分是由不同的地形組成的海洋。在上泥盆紀這些地區與北美-北歐-俄羅斯組成的大陸已開始有接觸。
顯生宙
石炭紀的發展
基本信息
- 中文名
- 顯生宙
- 外文名
- Phanerozoic Eon
- 概念
- 指“看得見生物的年代”
- 特點
- 多次地殼運動和氣候變化